Philander Brisson, 1762: Grey Four-eyed Opossums
Six species, one in Paraguay. Synonyms adapted from Gardner (2007).
Synonyms:
Didelphis Linnaeus 1758:54. In part.
Didelphys Schreber 1777:532. Unjustified emendation.
Philander Brisson 1762:13. Type species Didelphis oposum Linnaeus (1758).
Philander Tiedemann 1808:426. Type species Philander virginianus (=Didelphis oposum Linnaeus 1758)
Sarigua Muirhead 1819:429. In part.
Metachirus Burmeister 1854:135. In part. Described as a subgenus of Didelphis Linnaeus.
Gamba Liais 1872:329. In part.
Zygolestes Ameghino 1899:7. In part.
Metachirops Matschie 1916:262. Type species Didelphis quica Temminck (1824).
Holothylax Cabrera 1919:47. Type species Didelphis oposum Linnaeus (1758).
Metacherius Sanderson 1949:787. Incorrect spelling.
Phillander Rivillas, Caro, Caravajal & Vélez 2004:591. Incorrect spelling.
General characteristics: Medium to large grey or black opossums with conspicuous pale spots above the eye and in front of the ears. Ears naked. Tails exceeds the head and body length and is mostly naked, though furred at the base. Mature females have a well-developed pouch. Largely terrestrial forest inhabitants, though they do climb well. Fossils are known from the late Pliocene of Argentina and late Quaternary of Brazil.
Cranial characteristics: Similar to Didelphis in many respects but nasals less expanded laterally at maxillofrontal junction. Rostrum long and slender with broadly flared zygomatic arches. Postorbital constriction is smoothly rounded and narrow and temporal ridges converge to form a well-developed sagittal crest.
Genetic characteristics: 2n=14. The nucleolar organizing region (NOR) is located on two pairs of autosomes.
Parasites: Linardi (2006) gives the following ectoparasites recorded in this genus from Brazil: Siphanaptera: Craneopsylla (Stephanocircidae); Adoratopsylla (Ctenophthalmidae). Acari: Metastigmata Ixodes and Amblyomma (Ixodidae). Thatcher (2006) documents the following endoparasites for Brazil: Protozoa Trypanosoma. Trematoda Aphimerus (Opisthorchiidae), Rhopalias (Rhopaliasidae) and Paragonimus (Paragonimidae). Hershkovitz (1997) adds the following: Coleoptera Amblyopinus; Siphanaptera Ctenocephalides and Xenopsylla (Pulicidae) and Rhopalopsyllus (Rhopalopsyllidae); Nematoda Viannaia conspicua (Trichostrongylidae).
Taxonomy: Species limits in this genus are far from defined and though several geographically isolated species formerly included within the widespread Philander opossum have been recognised in recent years.
Paraguayan Species:
Philander quica - Southeastern Four-eyed Opossum HBook 9
Philander canus - Grey Four-eyed Opossum
Tribe Metachirini Reig, Kirsch & Marshall, 1985
General characteristics: Single large opossum with total length >500mm. Pouch absent.
Metachirus Burmeister, 1854: Brown Four-eyed Opossum
This is a monotypic genus. Synonyms adapted from Gardner (2007).
Synonyms:
Didelphis E. Geoffroy St.Hilaire 1803:142. Not Didelphis Linnaeus (1758).
Philander Gray 1843:100. In part. Not Philander Brisson (1762), Philander Beckmann (1772) or Philander Tiedemann (1808).
Metachirus Burmeister 1854:135. Type species Didelphis myosurus (=Didelphois myosurus Temminck 1824 =Didelphis nudicaudata E. Geoffroy St.Hilaire 1803). Described as a subgenus of Didelphis Linnaeus (1758).
Cuica Liais 1872:330. In part.
Lutreolina Bertoni 1939:6. Not Lutreolina O.Thomas 1910.
Philander Pine 1973:391. Not Philander Brisson (1762), Philander Beckmann (1772) or Philander Tiedemann (1808).
General characteristics: Dorsal pelage brownish with conspicuous pale spots above each eye and behind the ears. Ventrally paler. Sparsely-haired tail longer than the head and body lacks pigment on the terminal third.
Cranial characteristics: Skull slender with broad rostrum. Older individuals with elevated supraorbital ridges extending posteriorly to the parietals but not meeting along the midline. Palate with large and conspicuous maxillopalatine fenestrae. Maxillary and palatine fenestrae are absent. Bullae small, widely-separated and incompletely ossified.
Dental characteristics: Upper canines well-developed, though they are less than twice the height of P3. Lower canines shorter than p2. Molar series fairly long.
Genetic characteristics: 2n = 14 Karyotype morphologically similar to the ancestral marsupial complement and can be derived by as few as four changes in centromere position. The nucleolar organizing region (NOR) is located on a single pair of autosomes.
Parasites: Linardi (2006) gives the following ectoparasites recorded in this genus from Brazil: Siphanaptera: Adoratopsylla (Ctenophthalmidae). Acari: Astigmata Didelphoecius and Metachiroecius (Atopomelidae). Thatcher (2006) documents the following endoparasites for Brazil: Trematoda Podospathalium (Alariidae), Rhopalias (Rhopaliasidae). Cestoda Linstowia and Mathevotaenia (Anoplocephalidae). Nematoda Aspidodera (Aspidoderidae); Cruzia (Cruziidae); Physaloptera (Physalopteridae); Travassostrongylus and Viannaia (Trichostrongylidae). Acanthocephala Gigantorhynchus (Gigantorhynchidae).
Taxonomy: Patton & Costa (2003) found considerable molecular divergence in specimens from across the wide range of this currently monotypic genus, consistent with the existence of more than one species. Work remains to be done to define these species limits.
Paraguayan Species:
Metachirus nudicaudatus - Brown Four-eyed Opossum
Subfamily Thylamyinae Hershkovitz, 1992
General characteristics: Four species in three genera and two tribes. Pouch present and large number of teats.
Tribe Marmosopsini
Gracilinanus Gardner & Creighton, 1989: Mouse Opossums
Six species, one in Paraguay. Synonyms adapted from Gardner (2007).
Synonyms:
Didelphis JA Wagner 1842:359. Not Didelphis Linnaeus (1758).
Grymaeomys Burmeister 1854:27. In part. Proposed as a subgenus of Didelphis Linnaeus (1758).
Marmosa O.Thomas 1898:455. Not Marmosa Gray (1821).
Marmosa O.Thomas 1909:379. Not Marmosa Gray (1821).
Marmosa O.Thomas 1910:502. Not Marmosa Gray (1821).
Marmosa Cabrera 1919:34. In part. Not Marmosa Gray (1821).
Marmosa Tate 1931:10. Not Marmosa Gray (1821).
Marmosa Miranda-Ribeiro 1936:373. Not Marmosa Gray (1821).
Thylamys Miranda-Ribeiro 1936:387. Not Thylamys Gray (1843).
Marmosa Moojen 1943:2. In part. Not Marmosa Gray (1821).
Thylamys Cabrera 1958:26. In part. Not Thylamys Gray (1843).
Tylamys Ávila-Pires 1968:167. Incorrect spelling. Not Thylamys Gray (1843).
Gracilinanus Gardner & Creighton 1989:4. Type species Didelphys microtarsus JA Wagner (1842) by original designation.
General characteristics: Very small arboreal opossums with the ratio of tail to head and body length >1.3 but usually <1.5. Tail scales square or rounded up to 40 per centimetre, and arranged in rows, not in a spiral. Large ears. Tail weakly bicoloured in the Paraguayan species. Claws of the hands do not extend beyond the digital pads. Soles of the hand and feet are smooth and lack a granular appearance.
Cranial characteristics: Postorbital processes absent, supraorbital margin of the frontals often beaded in older specimens. Lamboidal crest weakly-developed or absent in older individuals. Hard palate highly fenestrated generally with three pairs of fenestrae - maxillary, maxillopalatine and palatine. Posterolateral palatal foramina are moderately-sized, approximately one third to half the width of M4 in length. Nasals expanded laterally at the maxillofrontal suture. Auditory bullae are large compared to related genera, but smaller proportionately than in Thylamys. Alisphenoid portion of the auditory bullae possesses a slender anteromedial process. Petrosal usually exposed between the squamosal and parietal.
Dental characteristics: Upper incisors increase in size from I2 through I5. Lower canines short. P2 equal in size to P3.
Genetic characteristics: 2n = 14 Karyotype morphologically similar to the ancestral marsupial complement and can be derived by as few as four changes in centromere position. The nucleolar organizing region (NOR) is located on a single pair of autosomes.
Parasites: Linardi (2006) gives the following ectoparasites recorded in this genus from Brazil: Acari: Metastigmata Ixodes and Amblyomma (Ixodidae). Acari: Mesostigmata Bdellonyssus (Macronyssidae). Acari: Astigmata Didelphoecius (Atopomelidae).
Paraguayan Species:
Gracilinanus agilis - Agile Gracile Opossum
Cryptonanus Voss, Lund & Jansa 2005: Cryptic Mouse Opossums
Five species, two in Paraguay. Synonyms adapted from Gardner (2007).
Grymaeomys Winge 1893:27. In part. Not Grymaeomys Burmeister (1854).
Marmosa Tate 1931:10. In part. Not Marmosa Gray (1821).
Marmosa Moojen 1943:2. In part. Not Marmosa Gray (1821).
Marmosa (Thylamys) Cabrera 1958:26. In part.
Gracilinanus Gardner & Creighton 1989:5. In part.
Gracilinanus Díaz, Flores & Barquez 2002:825. Not Gracilinanus Gardner & Creighton (1989).
Cryptonanus Voss, Lund & Jansa 2005:5. Type species Cryptonanus chacoensis (Tate 1931; originally Marmosa agilis chacoensis) by original description.
General characteristics: Outwardly extremely similar to Gracilinanus, in fact the Paraguayan representative of the genus is a cryptic species that was only recently distinguished from Gracilinanus agilis. These are extremely small opossums with a total length <260mm. The main distinguishing features refer to cranial characters and dentition.
Cranial characteristics: Maxillary fenestrae absent or extremely small. Projection of premaxillaries anterior to the incisors is lacking.
Dental characteristics: P2 smaller than P3.
Genetic characteristics: 2n = 14 Karyotype morphologically similar to the ancestral marsupial complement and can be derived by as few as four changes in centromere position. The nucleolar organizing region (NOR) is located on a single pair of autosomes.
Paraguayan Species:
Cryptonanus chacoensis - Chaco Mouse Opossum
Cryptonanus unduaviensis - Unduave Mouse Opossum
TribeThylamyini Hershkovitz, 1992
Thylamys Gray, 1843: Fat-tailed Opossums
Nine species, two in Paraguay. Synonyms adapted from Gardner (2007).
Synonyms:
Didelphis Desmarest 1804:19. In part. Not Didelphis Linnaeus (1758).
Didelphys Olfers 1818:205. Incorrect spelling. Not Didelphis Linnaeus (1758).
Sarigua Muirhead 1819:429. In part.
Didelphis Desmarest 1827:398. In part. Not Didelphis Linnaeus (1758).
Didelphys Waterhouse 1841:106. Incorrect spelling. Not Didelphis Linnaeus (1758).
Micoureus Lesson 1842:186. In part.
Didelphys JA Wagner 1842:360. Incorrect spelling. Not Didelphis Linnaeus (1758).
Thylamys Gray 1843:101. Type species Didelphis elegans Waterhouse (1839) by monotypy.
Didelphis Reinhardt 1851:v. In part. Not Didelphis Linnaeus (1758).
Grymaeomys Burmeister 1854:130. In part.
Microdelphys Burmeister 1856:86. In part. Proposed as a subgenus of Didelphis Linnaeus (1758).
Cuica Liais 1872:330. In part.
Didelphys (Micoureus) O.Thomas 1888:340. In part. Incorrect spelling. Not Didelphis Linnaeus (1758). Not Micoureus Lesson (1842).
Philander Cope 1889:130. In part. Not Philander Brisson (1762).
Didelphys RA Philippi 1894:36. Incorrect spelling. Not Didelphis Linnaeus (1758).
Micoureus Goeldi 1894:462. Not Micoureus Lesson (1842).
Marmosa O.Thomas 1894:188. In part. Not Marmosa Gray (1821).
Marmosa O.Thomas 1896:313 In part. Not Marmosa Gray (1821).
[Didelphys] (Peramys) Trouessart 1898:1244. In part. Incorrect spelling. Not Didelphis Linnaeus (1758). Not Peramys Lesson (1842).
Marmosa O.Thomas 1902:158. Not Marmosa Gray (1821).
Marmosa O.Thomas 1912:409. Not Marmosa Gray (1821).
Didelphis (Dromicops) Matschie 1916:271. Not Didelphis Linnaeus (1758). Not Dromicops O.Thomas (1894).
Marmosa O.Thomas 1921a:186. Not Marmosa Gray (1821).
Marmosa O.Thomas 1921b:519. Not Marmosa Gray (1821).
Marmosa O.Thomas 1926:327. Not Marmosa Gray (1821).
Marmosa Tate 1931:14. Not Marmosa Gray (1821).
Marmosa Marelli 1932:68. Not Marmosa Gray (1821).
Thylamis A.Miranda-Ribeiro 1936:328. Incorrect spelling.
Marmosa Handley 1957:402. Not Marmosa Gray (1821).
Didelphys (Paramys) Cabrera 1958:29. Incorrect spelling. Not Peramys Lesson (1842).
Dromictops Cabrera 1958:30. Incorrect spelling. Not Dromicops O.Thomas (1894).
Marmosa Petter 1968:313. Not Marmosa Gray (1821).
Thulamys Reig, Kirsch & Marshall 1985:336. Incorrect spelling.
Macrodelphys BE Brown 2004:145. Incorrect spelling.
General characteristics: Small opossums with the ratio of tail to head and body length in the region of 1 to 1.25. Bilaterally symmetrical rows of teats (as opposed to teats arranged in a circular pattern as in other genera) include inguinal, abdominal and pectoral teats and number 15 to 19 (7-1-7 or rarely 9-1-9). Feet are small and white with short digits and well-developed ungual tufts reaching the claw tips. Claws on the forefoot digits II-IV extend well beyond the digital pads. Central soles of the feet are covered with small granular tubercles. Thenar and hypothenar pads are not fused with interdigitals. The tail is bicoloured in many species and at certain times of year some species store fat in the basal section of their tails. Tail scales are tiny (35 or more rows per cm), rounded or square in shape and arranged in rings. Distribution of vibrissae (per side of body): 2 superciliary, 5 to 8 genal, 3 submental, 2 interramal, 1 antebrachial, 1 anconeal, and 3 to 4 carpal. Fossil species of this genus are known from the late Pleistocene.
Cranial characteristics: Postorbital processes usually absent, but may appear as ridges or postorbital projections in adults of some species (eg. T.pusillus). Nasals are elongated, slender and only slightly expanded at the maxillofrontal suture. Auditory bullae are large, rounded and not widely separated - ratio of distance between bullae to width of a single bulla is 1.5 or less. An anteromedial alisphenoid process is present. Palate may have 2 or 3 paired fenestrae. Posterolateral palatal foramina are extremely long and the width of each is equal to or greater than the width of the last molar.
Dental characteristics: P3 exceeds P2 in height and anteroposterior length. Molars wide and compressed lengthways. Paracone of M2 slightly displaced towards lingual border. Lower canines with well-developed cusp.
Genetic characteristics: 2n = 14 Karyotype morphologically similar to the ancestral marsupial complement and can be derived by as few as four changes in centromere position. The nucleolar organizing region (NOR) is located on a single pair of autosomes.
Parasites: Linardi (2006) gives the following ectoparasite recorded in this genus from Brazil: Acari: Astigmata Didelphoecius (Atopomelidae).
Taxonomy: The historical taxonomy of this genus is summarised by Solari (2003).
Paraguayan Species:
Thylamys macrurus - Long-tailed Fat-tailed Opossum
Thylamys pusillus - Chaco Fat-tailed Opossum
Subfamily Marmosinae Reig, Kirsch & Marshall, 1985
General characteristics: Five species in two genera and two tribes. Pouch absent and large number of teats.
Tribe Marmosini Reig, Kirsch & Marshall, 1985
Subgenus Micoureus Lesson, 1842: Woolly Mouse Opossums
Six species, two in Paraguay. Synonyms adapted from Gardner (2007).
Synonyms:
Didelphis Temminck 1824:46. In part. Not Didelphis Linnaeus (1758).
Micoureus Lesson 1842:186. Type species Didelphis cinerea Temminck (1824) by subsequent designation.
Philander Gray 1843:101. In part. Not Philander Brisson (1762), Philander Beckmann (1772) or Philander Tiedemann (1808).
[Didelphys (]Marmosa [)] Trouessart 1905:855. In part. Incorrect spelling. Not Marmosa Gray (1821).
Caluromys Matschie 1916:269. Not Caluromys JA Allen (1900).
Micoures Reig, Kirsch & Marshall 1985:337. Incorrect spelling.
Micoures Massoia 1988:6. Incorrect spelling.
General characteristics: Medium-sized Didelphids with tail >1.3x head and body length. Dorsal pelage is long, thick and woolly, ventral pelage is smoother and softer. Mammae are abdominal or inguinal and vary in number from 9 to 15. Tail scales are coarse and rhomboid, arranged in a spiral with 14 to 16 rows per centimetre. Fur extends approximately 5cm onto the base of the tail. Interscalar hairs of tail are slender, of approximately equal length and thickness and occur in triplets under the posterior margin of each scale. Distal third of the tail is white in the Paraguayan species, though this is not typical of the genus. Feet are broad with the claws of the forefeet extending slightly beyond the digital pads. Thenar and first interdigital pads are fused on the hindfoot but lie together on the forefoot. Fourth interdigital pad lies against the hypothenar pad of the forefoot but the two are either fused or in direct contact on the hindfoot. Central part of the soles of all feet are smooth. Digit IV on the hindfoot is longest with a length ratio of 0.45 when compared to the hindfoot length. Second and third interdigital pads on all feet are triangular and approximately as wide as they are long. Ventral surfaces of the digits have transverse bars. Fossil species of this genus are known from the late Pleistocene.
Cranial characteristics: Skull large with broad zygomatic arches. Postorbital processes are well-developed and beaded. Nasals abruptly expanded at the maxillofrontal suture. Lacks sagittal crest, temporal ridges being subparallel or convergent posteriorly. Prominent lamboidal crest. Auditory bullae large and set wide apart - the ratio of the distance between the bullae to the width of one bulla >1.5. Alisphenoid component of the bulla is hemispherical and lacking an anteromedial process. Ectotympanic expands laterally forming the ventral wall of the bulla. Maxillary fenestrae absent or extremely small. Maxillopalatine fenestrae and posterolateral foramina present, latter approximately equal to half the width of the last upper molar. Projection of premaxillaries anterior to the incisors is lacking.
Dental characteristics: I1 is longest and separated from I2 by a space. Incisor length increases from I2 through to I5. P2 is larger than P3 and M3 is the widest upper molar. Canines are long and curved.
Skeletal characteristics: Caudal vertebrae vary in number from 32-35.
Genetic characteristics: 2n = 14 Karyotype morphologically similar to the ancestral marsupial complement and can be derived by as few as four changes in centromere position. The nucleolar organizing region (NOR) is located on two pairs of autosomes.
Parasites: Linardi (2006) gives the following ectoparasites recorded in this genus from Brazil: Siphanaptera: Adoratopsylla (Ctenophthalmidae). Acari: Mesostigmata Bdellonyssus (Macronyssidae). Acari: Astigmata Didelphoecius (Atopomelidae).
Paraguayan Species:
Marmosa (Micoureus) constantiae - White-bellied Woolly Mouse Opossum
Marmosa (Micoureus) paraguayana - Long-furred Woolly Mouse Opossum
Tribe Monodelphini Hershkovitz, 1992
Monodelphis Burnett, 1830: Short-tailed Opossums
Twenty-nine species, three present in Paraguay.
Synonyms:
Sorex Müller 1776:36. In part. Not Sorex Linnaeus (1758).
Didelphis Erxleben 1777:80. In part. Not Didelphis Linnaeus (1758).
Didelphys Schreber 1777:549. In part. Unjustified emendation.
Viverra G.Shaw 1800:432. In part. Not Viverra Linnaeus (1758).
Mustela Daudin in Lacépède 1802:163. In part. Not Mustela Linnaeus (1758).
Philander Tiedemann 1808:428. In part.
Sarigua Muirhead 1819:429. In part.
Monodelphis Burnett 1830:351. Type species Monodelphis Brachyura Burnett (1830) (=Didelphys brachyuros Schreber 1777 [=Didelphis brevicaudata Erxleben 1777]) by selection.
? Crossopus Lesson 1842:91. In part. Not Crossopus Wagler (1832).
Micoureus Lesson 1842:186. In part.
Peramys Lesson 1842:187. Type species Peramys brachyurus Lesson (1842) (=Didelphis dimidiata JA Wagner (1847) by selection. Not Didelphis brevicaudata Erxleben (1777).
Grymaeomys Burmeister 1854:138. In part.
Hemiurus P.Gervais 1855:287. Preoccupied.
Microdelphys Burmeister 1856:83. Type species Didelphis (Microdelphys) tristriata Burmeister (1856) (=Didelphys tristriata Illiger 1815, [=Sorex americanus Müller 1776]) by designation. Proposed as a subgenus of Didelphis Linnaeus (1758).
Microdidelphys Trouessart 1898:1242. Incorrect spelling.
Monodelphiops Matschie 1916:261. Type species Microdelphys sorex Hensel (1872) by designation.
Minuania Cabrera 1919:30. Type species Didelphis dimidiata JA Wagner (1847) by description.
Microdelphis Pohle 1927:240. Incorrect spelling.
Monodelphys Reig 1959:57. Incorrect spelling.
Monodelhpis Tálice, Lafitte de Mosera & Machado 1960:151. Incorrect spelling.
General characteristics: Three species of small, shrew-like opossums, with very short tails - 50-70% of the head and body length. Species are extremely similar and distinguished with care, all having short, stout legs, short ears, long-pointed snouts continuous with a steep forehead and a short prehensile-tipped tail. Toes are short, the fifth digit of the hind foot being particularly so and not opposable with the rest of the foot. Claws are long and some species show modifications of the forefeet for digging. Foot pads are poorly developed. Pelage is never long and lax, but varies from short and smooth to long and woolly and guard hairs are absent. Predominately terrestrial and crepuscular in habits, the life cycle is short and neither sex survives to reproduce a second season. No fossil remains have been found that can be assigned to the Pleistocene or earlier.
Cranial characteristics: Palate lacks a second posterior pair of vacuities. Posterolateral foramina reduced. Poorly developed orbitae and lachrimal foramina opening outside the orbit. Alisphenoid with lateral bony rib. Paraoccipital processes well-developed.
Dental characteristics: First premolars not greatly reduced. Third premolar larger or much larger than second premolar on upper and lower jaws. Premolars typically increase in size from front to back. Canine fossae well-developed. Upper first incisor smaller than I2-I4. M4 with vestigial metacone. Lower third premolar larger than p2. m1-4 with short talonids. Poorly developed hypoconids and entoconids. m4 with well-developed cingulum labial to its talonid.
Genetic characteristics: 2n=18. Karyotype can be readily derived from the ancestral marsupial 2n = 14 complement by two fissions and several minor changes in centromere position. NORs have been reported from the short arms of chromosome 5 and the X.
Parasites: This genus shares many ectoparasites with rodents which constitute a major part of the diet. Linardi (2006) gives the following ectoparasites recorded in this genus from Brazil: Siphanaptera: Craneopsylla (Stephanocircidae); Polygenis (Rhopalopsyllidae); Adoratopsylla (Ctenophthalmidae). Acari: Metastigmata Ixodes, Ornithodoros and Amblyomma (Ixodidae). Acari: Mesostigmata Ornithonyssus and Lepronyssoides (Macronyssidae); Eubrachylaelaps, Laelaps Mysolaelaps and Gigantolaelaps (Laelapidae). Acari: Prostigmata Parasecia and Trombewingi (Trombiculidae). Thatcher (2006) documents the following endoparasite for Brazil: Cestoda Linstowia (Anoplocephalidae).
Paraguayan Species:
Monodelphis domestica - Grey Short-tailed Opossum
Monodelphis kunsi - Pygmy Short-tailed Opossum
Monodelphis dimidiata - Red-sided Short-tailed Opossum
Two further species of Monodelphis have been stated to occur in Paraguay in the published literature. M.scalops (Contreras & Silveira 1995) was shown to be an incorrectly identified specimen of M.sorex (de la Sancha et al 2007). Monodelphis brevicaudis mapped for the Paraguayan Chaco by Brown (2004) has variously been regarded to be a senior synonym of M.domestica or M.brevicaudata, but is now generally considered a synonym of M.sorex (de la Sancha et al 2007). Other species may occur in Paraguay but their presence is yet to be proven.
Key to Adults of the Paraguayan Species of Opossum
The fifteen species documented as occurring in Paraguay are keyed as well as three further species considered of hypothetical occurrence: Philander opossum, Micoureus constantiae and Gracilinanus microtarsus.
1a Pelage long and woolly, long prehensile tail furred along basal third, conspicuous medial facial stripe, large forward-facing eyes and arboreal behaviour.................................................................Caluromys lanatus
1b Not as above.....................................................................................................................................................2
2a Size small to medium, terrestrial and typically diurnal in behaviour. Aspect shrew-like or mouse-like with short tail <70% of head and body length........................................................................................... Monodelphis 3
2b Not as above....................................................................................................................................................5
3a Colouration uniform on dorsum without contrasting colouration on flanks, head or rump............................. 4
3b Flanks reddish, contrasting with browner dorsum and paler underparts. Rump dark reddish. Fur short and smooth, lying flat against body....................................................................................Monodelphis dimidiata
4a Size small (hindfoot with claw <16mm). Dorsal colouration uniformly reddish or reddish-brown..........................................................................................................................Monodelphis kunsi
4b Size larger (hindfoot with claw >18mm). Dorsal colouration uniformly grey or grey-brown.....................................................................................................................Monodelphis domestica
5a Conspciuous pale spots above eye and at base of ears..................................................................................... 6
5b No conspicuous pale spots above eye and at base of ears................................................................................8
6a Dorsal pelage predominately brown. Secondary pale spots large and located behind ears. Tail naked at base and uniformly-coloured or paling gradually towards tip - not sharply bicoloured. Female lacks pouch.................................................... ........................................................... Metachirus nudicaudatus
6b Dorsal pelage short and predominately greyish or blackish. Secondary pale spots small and located in front of ears. Tail furred for basal 6-8cm and sharply bicoloured. Pouch present in female................................Philander 7
7a Incomplete labial cingulum on P3. Atlantic Forest............................................................................................................................Philander quica
7b Complete labial cingulum on P3. Chaco and Paraguay River Basin. ...........................................................................................................................................Philander canus
8a Dorsal pelage dense and velvety, predominately black with grey markings. Tail furred for basal 10%, naked and black for rest of length with whitish tip. Webbed feet. Swims well......................Chironectes minimus
8b Not as above....................................................................................................................................................9
9a Ears extremely short, rounded and dark, barely projecting above fur. General appearance weasel-like. Pelage uniformly brownish or reddish. Tail extremely thick at base, and furred along basal third. Mainly terrestrial but climbs and swims readily.....................................................................................Lutreolina crassicaudata
9b Not as above...................................................................................................................................................10
10a Size large (total length >500mm). Pelage with long, prominent guard hairs and soft, dense underfur. Females with well-developed marsupium..............................................................................................Didelphis 11
10b Size small or medium (total length generally much less than 450mm) . General appearance mouse-like. Pouch absent........................................................................................................................................................12
11a Dorsal pelage mostly greyish. Prominent white facial markings. Ears pale. Widespread in a variety of habitats.......................................................................................................................Didelphis albiventris
11b Dorsal pelage mostly blackish. Golden-buff facial markings. Ears dark. Forested areas only................................................................................................................................Didelphis aurita
12a Size medium (TL>350mm), white tip to tail..................................................................................Micoureus 13
12b Size small (TL <200mm). Pelage short. Usually no white tip to tail.............................................................14
13a Fur long and woolly extending 30mm or so down base of tail. Ventral pelage buffy to cinnamon and grey-based. Southern and eastern Paraguay in Atlantic Forest....................................Micoureus paraguayanus
13b Fur long but not overly dense and not notably extending onto base of tail. Ventral pelage creamy to yellow-white and white-based. In Chaco and Cerrado.....................Micoureus constantiae
14a Ratio of tail length to head and body length between 1 and 1.25. Fat may be stored at base of tail at certain times of year. Soles of feet densely covered with small tubercles. Length of digit IV <0.45 times the hindfoot length. Teats arranged in bilaterally symmetrical rows...........................................................................Thylamys 15
14b Ratio of tail length to head and body length between 1 and 1.96. Fat never stored at base of tail. Tail scales rounded or square with annular arrangement. Soles of feet smooth or sparsely granulated. Length of digit IV >0.45 times the hindfoot length. Teats with typical circular arrangement..............................................................16
15a Size large, tail >135mm. Weight >40g. Tail with whitish tip. Orient only...........Thylamys macrurus
15b Size small, tail <135mm. Weight <35g. Tail lacks whitish tip. Chaco only...........Thylamys pusillus
16a Ventral pelage creamy with greyish base. Ratio of tail length to head and body length >1.2 but usually <1.5. Tail length 110-165mm. P2 and P3 of equal height..........................................................................Gracilinanus 17
16b Ventral pelage not grey-based. Ratio of tail length to head and body length <1.2. Tail length 95-117mm. P2 smaller than P3......................................................................................................Cryptonanus chacoensis / unduaviensis.
17a Dorsal pelage pale brown to greyish-brown looking grizzled. Ventral pelage grey-based except for chin, throat, upper breast and scrotal area. Ocular patch does not extend to nose and ears. Face not markedly paler than dorsum. Tail typically <140mm. Ears usually >21mm. Mainly cerrado in distribution..................................................................................................................Gracilinanus agilis
17b Dorsal pelage reddish-brown to chestnut-brown looking uniform. Ventral entirely grey-based except for chin. Ocular patch extends to nose and ears. Face contrastingly paler than dorsum. Tail typically >140mm. Ears usually <21mm. Hypothetical in Atlantic Forest..................................................................Gracilinanus microtarsus
REFERENCES
Brown BE 2004 - Atlas of New World Marsupials - Fieldiana Zoology 1257.
Six species, one in Paraguay. Synonyms adapted from Gardner (2007).
Synonyms:
Didelphis Linnaeus 1758:54. In part.
Didelphys Schreber 1777:532. Unjustified emendation.
Philander Brisson 1762:13. Type species Didelphis oposum Linnaeus (1758).
Philander Tiedemann 1808:426. Type species Philander virginianus (=Didelphis oposum Linnaeus 1758)
Sarigua Muirhead 1819:429. In part.
Metachirus Burmeister 1854:135. In part. Described as a subgenus of Didelphis Linnaeus.
Gamba Liais 1872:329. In part.
Zygolestes Ameghino 1899:7. In part.
Metachirops Matschie 1916:262. Type species Didelphis quica Temminck (1824).
Holothylax Cabrera 1919:47. Type species Didelphis oposum Linnaeus (1758).
Metacherius Sanderson 1949:787. Incorrect spelling.
Phillander Rivillas, Caro, Caravajal & Vélez 2004:591. Incorrect spelling.
General characteristics: Medium to large grey or black opossums with conspicuous pale spots above the eye and in front of the ears. Ears naked. Tails exceeds the head and body length and is mostly naked, though furred at the base. Mature females have a well-developed pouch. Largely terrestrial forest inhabitants, though they do climb well. Fossils are known from the late Pliocene of Argentina and late Quaternary of Brazil.
Cranial characteristics: Similar to Didelphis in many respects but nasals less expanded laterally at maxillofrontal junction. Rostrum long and slender with broadly flared zygomatic arches. Postorbital constriction is smoothly rounded and narrow and temporal ridges converge to form a well-developed sagittal crest.
Genetic characteristics: 2n=14. The nucleolar organizing region (NOR) is located on two pairs of autosomes.
Parasites: Linardi (2006) gives the following ectoparasites recorded in this genus from Brazil: Siphanaptera: Craneopsylla (Stephanocircidae); Adoratopsylla (Ctenophthalmidae). Acari: Metastigmata Ixodes and Amblyomma (Ixodidae). Thatcher (2006) documents the following endoparasites for Brazil: Protozoa Trypanosoma. Trematoda Aphimerus (Opisthorchiidae), Rhopalias (Rhopaliasidae) and Paragonimus (Paragonimidae). Hershkovitz (1997) adds the following: Coleoptera Amblyopinus; Siphanaptera Ctenocephalides and Xenopsylla (Pulicidae) and Rhopalopsyllus (Rhopalopsyllidae); Nematoda Viannaia conspicua (Trichostrongylidae).
Taxonomy: Species limits in this genus are far from defined and though several geographically isolated species formerly included within the widespread Philander opossum have been recognised in recent years.
Paraguayan Species:
Philander quica - Southeastern Four-eyed Opossum HBook 9
Philander canus - Grey Four-eyed Opossum
Tribe Metachirini Reig, Kirsch & Marshall, 1985
General characteristics: Single large opossum with total length >500mm. Pouch absent.
Metachirus Burmeister, 1854: Brown Four-eyed Opossum
This is a monotypic genus. Synonyms adapted from Gardner (2007).
Synonyms:
Didelphis E. Geoffroy St.Hilaire 1803:142. Not Didelphis Linnaeus (1758).
Philander Gray 1843:100. In part. Not Philander Brisson (1762), Philander Beckmann (1772) or Philander Tiedemann (1808).
Metachirus Burmeister 1854:135. Type species Didelphis myosurus (=Didelphois myosurus Temminck 1824 =Didelphis nudicaudata E. Geoffroy St.Hilaire 1803). Described as a subgenus of Didelphis Linnaeus (1758).
Cuica Liais 1872:330. In part.
Lutreolina Bertoni 1939:6. Not Lutreolina O.Thomas 1910.
Philander Pine 1973:391. Not Philander Brisson (1762), Philander Beckmann (1772) or Philander Tiedemann (1808).
General characteristics: Dorsal pelage brownish with conspicuous pale spots above each eye and behind the ears. Ventrally paler. Sparsely-haired tail longer than the head and body lacks pigment on the terminal third.
Cranial characteristics: Skull slender with broad rostrum. Older individuals with elevated supraorbital ridges extending posteriorly to the parietals but not meeting along the midline. Palate with large and conspicuous maxillopalatine fenestrae. Maxillary and palatine fenestrae are absent. Bullae small, widely-separated and incompletely ossified.
Dental characteristics: Upper canines well-developed, though they are less than twice the height of P3. Lower canines shorter than p2. Molar series fairly long.
Genetic characteristics: 2n = 14 Karyotype morphologically similar to the ancestral marsupial complement and can be derived by as few as four changes in centromere position. The nucleolar organizing region (NOR) is located on a single pair of autosomes.
Parasites: Linardi (2006) gives the following ectoparasites recorded in this genus from Brazil: Siphanaptera: Adoratopsylla (Ctenophthalmidae). Acari: Astigmata Didelphoecius and Metachiroecius (Atopomelidae). Thatcher (2006) documents the following endoparasites for Brazil: Trematoda Podospathalium (Alariidae), Rhopalias (Rhopaliasidae). Cestoda Linstowia and Mathevotaenia (Anoplocephalidae). Nematoda Aspidodera (Aspidoderidae); Cruzia (Cruziidae); Physaloptera (Physalopteridae); Travassostrongylus and Viannaia (Trichostrongylidae). Acanthocephala Gigantorhynchus (Gigantorhynchidae).
Taxonomy: Patton & Costa (2003) found considerable molecular divergence in specimens from across the wide range of this currently monotypic genus, consistent with the existence of more than one species. Work remains to be done to define these species limits.
Paraguayan Species:
Metachirus nudicaudatus - Brown Four-eyed Opossum
Subfamily Thylamyinae Hershkovitz, 1992
General characteristics: Four species in three genera and two tribes. Pouch present and large number of teats.
Tribe Marmosopsini
Gracilinanus Gardner & Creighton, 1989: Mouse Opossums
Six species, one in Paraguay. Synonyms adapted from Gardner (2007).
Synonyms:
Didelphis JA Wagner 1842:359. Not Didelphis Linnaeus (1758).
Grymaeomys Burmeister 1854:27. In part. Proposed as a subgenus of Didelphis Linnaeus (1758).
Marmosa O.Thomas 1898:455. Not Marmosa Gray (1821).
Marmosa O.Thomas 1909:379. Not Marmosa Gray (1821).
Marmosa O.Thomas 1910:502. Not Marmosa Gray (1821).
Marmosa Cabrera 1919:34. In part. Not Marmosa Gray (1821).
Marmosa Tate 1931:10. Not Marmosa Gray (1821).
Marmosa Miranda-Ribeiro 1936:373. Not Marmosa Gray (1821).
Thylamys Miranda-Ribeiro 1936:387. Not Thylamys Gray (1843).
Marmosa Moojen 1943:2. In part. Not Marmosa Gray (1821).
Thylamys Cabrera 1958:26. In part. Not Thylamys Gray (1843).
Tylamys Ávila-Pires 1968:167. Incorrect spelling. Not Thylamys Gray (1843).
Gracilinanus Gardner & Creighton 1989:4. Type species Didelphys microtarsus JA Wagner (1842) by original designation.
General characteristics: Very small arboreal opossums with the ratio of tail to head and body length >1.3 but usually <1.5. Tail scales square or rounded up to 40 per centimetre, and arranged in rows, not in a spiral. Large ears. Tail weakly bicoloured in the Paraguayan species. Claws of the hands do not extend beyond the digital pads. Soles of the hand and feet are smooth and lack a granular appearance.
Cranial characteristics: Postorbital processes absent, supraorbital margin of the frontals often beaded in older specimens. Lamboidal crest weakly-developed or absent in older individuals. Hard palate highly fenestrated generally with three pairs of fenestrae - maxillary, maxillopalatine and palatine. Posterolateral palatal foramina are moderately-sized, approximately one third to half the width of M4 in length. Nasals expanded laterally at the maxillofrontal suture. Auditory bullae are large compared to related genera, but smaller proportionately than in Thylamys. Alisphenoid portion of the auditory bullae possesses a slender anteromedial process. Petrosal usually exposed between the squamosal and parietal.
Dental characteristics: Upper incisors increase in size from I2 through I5. Lower canines short. P2 equal in size to P3.
Genetic characteristics: 2n = 14 Karyotype morphologically similar to the ancestral marsupial complement and can be derived by as few as four changes in centromere position. The nucleolar organizing region (NOR) is located on a single pair of autosomes.
Parasites: Linardi (2006) gives the following ectoparasites recorded in this genus from Brazil: Acari: Metastigmata Ixodes and Amblyomma (Ixodidae). Acari: Mesostigmata Bdellonyssus (Macronyssidae). Acari: Astigmata Didelphoecius (Atopomelidae).
Paraguayan Species:
Gracilinanus agilis - Agile Gracile Opossum
Cryptonanus Voss, Lund & Jansa 2005: Cryptic Mouse Opossums
Five species, two in Paraguay. Synonyms adapted from Gardner (2007).
Grymaeomys Winge 1893:27. In part. Not Grymaeomys Burmeister (1854).
Marmosa Tate 1931:10. In part. Not Marmosa Gray (1821).
Marmosa Moojen 1943:2. In part. Not Marmosa Gray (1821).
Marmosa (Thylamys) Cabrera 1958:26. In part.
Gracilinanus Gardner & Creighton 1989:5. In part.
Gracilinanus Díaz, Flores & Barquez 2002:825. Not Gracilinanus Gardner & Creighton (1989).
Cryptonanus Voss, Lund & Jansa 2005:5. Type species Cryptonanus chacoensis (Tate 1931; originally Marmosa agilis chacoensis) by original description.
General characteristics: Outwardly extremely similar to Gracilinanus, in fact the Paraguayan representative of the genus is a cryptic species that was only recently distinguished from Gracilinanus agilis. These are extremely small opossums with a total length <260mm. The main distinguishing features refer to cranial characters and dentition.
Cranial characteristics: Maxillary fenestrae absent or extremely small. Projection of premaxillaries anterior to the incisors is lacking.
Dental characteristics: P2 smaller than P3.
Genetic characteristics: 2n = 14 Karyotype morphologically similar to the ancestral marsupial complement and can be derived by as few as four changes in centromere position. The nucleolar organizing region (NOR) is located on a single pair of autosomes.
Paraguayan Species:
Cryptonanus chacoensis - Chaco Mouse Opossum
Cryptonanus unduaviensis - Unduave Mouse Opossum
TribeThylamyini Hershkovitz, 1992
Thylamys Gray, 1843: Fat-tailed Opossums
Nine species, two in Paraguay. Synonyms adapted from Gardner (2007).
Synonyms:
Didelphis Desmarest 1804:19. In part. Not Didelphis Linnaeus (1758).
Didelphys Olfers 1818:205. Incorrect spelling. Not Didelphis Linnaeus (1758).
Sarigua Muirhead 1819:429. In part.
Didelphis Desmarest 1827:398. In part. Not Didelphis Linnaeus (1758).
Didelphys Waterhouse 1841:106. Incorrect spelling. Not Didelphis Linnaeus (1758).
Micoureus Lesson 1842:186. In part.
Didelphys JA Wagner 1842:360. Incorrect spelling. Not Didelphis Linnaeus (1758).
Thylamys Gray 1843:101. Type species Didelphis elegans Waterhouse (1839) by monotypy.
Didelphis Reinhardt 1851:v. In part. Not Didelphis Linnaeus (1758).
Grymaeomys Burmeister 1854:130. In part.
Microdelphys Burmeister 1856:86. In part. Proposed as a subgenus of Didelphis Linnaeus (1758).
Cuica Liais 1872:330. In part.
Didelphys (Micoureus) O.Thomas 1888:340. In part. Incorrect spelling. Not Didelphis Linnaeus (1758). Not Micoureus Lesson (1842).
Philander Cope 1889:130. In part. Not Philander Brisson (1762).
Didelphys RA Philippi 1894:36. Incorrect spelling. Not Didelphis Linnaeus (1758).
Micoureus Goeldi 1894:462. Not Micoureus Lesson (1842).
Marmosa O.Thomas 1894:188. In part. Not Marmosa Gray (1821).
Marmosa O.Thomas 1896:313 In part. Not Marmosa Gray (1821).
[Didelphys] (Peramys) Trouessart 1898:1244. In part. Incorrect spelling. Not Didelphis Linnaeus (1758). Not Peramys Lesson (1842).
Marmosa O.Thomas 1902:158. Not Marmosa Gray (1821).
Marmosa O.Thomas 1912:409. Not Marmosa Gray (1821).
Didelphis (Dromicops) Matschie 1916:271. Not Didelphis Linnaeus (1758). Not Dromicops O.Thomas (1894).
Marmosa O.Thomas 1921a:186. Not Marmosa Gray (1821).
Marmosa O.Thomas 1921b:519. Not Marmosa Gray (1821).
Marmosa O.Thomas 1926:327. Not Marmosa Gray (1821).
Marmosa Tate 1931:14. Not Marmosa Gray (1821).
Marmosa Marelli 1932:68. Not Marmosa Gray (1821).
Thylamis A.Miranda-Ribeiro 1936:328. Incorrect spelling.
Marmosa Handley 1957:402. Not Marmosa Gray (1821).
Didelphys (Paramys) Cabrera 1958:29. Incorrect spelling. Not Peramys Lesson (1842).
Dromictops Cabrera 1958:30. Incorrect spelling. Not Dromicops O.Thomas (1894).
Marmosa Petter 1968:313. Not Marmosa Gray (1821).
Thulamys Reig, Kirsch & Marshall 1985:336. Incorrect spelling.
Macrodelphys BE Brown 2004:145. Incorrect spelling.
General characteristics: Small opossums with the ratio of tail to head and body length in the region of 1 to 1.25. Bilaterally symmetrical rows of teats (as opposed to teats arranged in a circular pattern as in other genera) include inguinal, abdominal and pectoral teats and number 15 to 19 (7-1-7 or rarely 9-1-9). Feet are small and white with short digits and well-developed ungual tufts reaching the claw tips. Claws on the forefoot digits II-IV extend well beyond the digital pads. Central soles of the feet are covered with small granular tubercles. Thenar and hypothenar pads are not fused with interdigitals. The tail is bicoloured in many species and at certain times of year some species store fat in the basal section of their tails. Tail scales are tiny (35 or more rows per cm), rounded or square in shape and arranged in rings. Distribution of vibrissae (per side of body): 2 superciliary, 5 to 8 genal, 3 submental, 2 interramal, 1 antebrachial, 1 anconeal, and 3 to 4 carpal. Fossil species of this genus are known from the late Pleistocene.
Cranial characteristics: Postorbital processes usually absent, but may appear as ridges or postorbital projections in adults of some species (eg. T.pusillus). Nasals are elongated, slender and only slightly expanded at the maxillofrontal suture. Auditory bullae are large, rounded and not widely separated - ratio of distance between bullae to width of a single bulla is 1.5 or less. An anteromedial alisphenoid process is present. Palate may have 2 or 3 paired fenestrae. Posterolateral palatal foramina are extremely long and the width of each is equal to or greater than the width of the last molar.
Dental characteristics: P3 exceeds P2 in height and anteroposterior length. Molars wide and compressed lengthways. Paracone of M2 slightly displaced towards lingual border. Lower canines with well-developed cusp.
Genetic characteristics: 2n = 14 Karyotype morphologically similar to the ancestral marsupial complement and can be derived by as few as four changes in centromere position. The nucleolar organizing region (NOR) is located on a single pair of autosomes.
Parasites: Linardi (2006) gives the following ectoparasite recorded in this genus from Brazil: Acari: Astigmata Didelphoecius (Atopomelidae).
Taxonomy: The historical taxonomy of this genus is summarised by Solari (2003).
Paraguayan Species:
Thylamys macrurus - Long-tailed Fat-tailed Opossum
Thylamys pusillus - Chaco Fat-tailed Opossum
Subfamily Marmosinae Reig, Kirsch & Marshall, 1985
General characteristics: Five species in two genera and two tribes. Pouch absent and large number of teats.
Tribe Marmosini Reig, Kirsch & Marshall, 1985
Subgenus Micoureus Lesson, 1842: Woolly Mouse Opossums
Six species, two in Paraguay. Synonyms adapted from Gardner (2007).
Synonyms:
Didelphis Temminck 1824:46. In part. Not Didelphis Linnaeus (1758).
Micoureus Lesson 1842:186. Type species Didelphis cinerea Temminck (1824) by subsequent designation.
Philander Gray 1843:101. In part. Not Philander Brisson (1762), Philander Beckmann (1772) or Philander Tiedemann (1808).
[Didelphys (]Marmosa [)] Trouessart 1905:855. In part. Incorrect spelling. Not Marmosa Gray (1821).
Caluromys Matschie 1916:269. Not Caluromys JA Allen (1900).
Micoures Reig, Kirsch & Marshall 1985:337. Incorrect spelling.
Micoures Massoia 1988:6. Incorrect spelling.
General characteristics: Medium-sized Didelphids with tail >1.3x head and body length. Dorsal pelage is long, thick and woolly, ventral pelage is smoother and softer. Mammae are abdominal or inguinal and vary in number from 9 to 15. Tail scales are coarse and rhomboid, arranged in a spiral with 14 to 16 rows per centimetre. Fur extends approximately 5cm onto the base of the tail. Interscalar hairs of tail are slender, of approximately equal length and thickness and occur in triplets under the posterior margin of each scale. Distal third of the tail is white in the Paraguayan species, though this is not typical of the genus. Feet are broad with the claws of the forefeet extending slightly beyond the digital pads. Thenar and first interdigital pads are fused on the hindfoot but lie together on the forefoot. Fourth interdigital pad lies against the hypothenar pad of the forefoot but the two are either fused or in direct contact on the hindfoot. Central part of the soles of all feet are smooth. Digit IV on the hindfoot is longest with a length ratio of 0.45 when compared to the hindfoot length. Second and third interdigital pads on all feet are triangular and approximately as wide as they are long. Ventral surfaces of the digits have transverse bars. Fossil species of this genus are known from the late Pleistocene.
Cranial characteristics: Skull large with broad zygomatic arches. Postorbital processes are well-developed and beaded. Nasals abruptly expanded at the maxillofrontal suture. Lacks sagittal crest, temporal ridges being subparallel or convergent posteriorly. Prominent lamboidal crest. Auditory bullae large and set wide apart - the ratio of the distance between the bullae to the width of one bulla >1.5. Alisphenoid component of the bulla is hemispherical and lacking an anteromedial process. Ectotympanic expands laterally forming the ventral wall of the bulla. Maxillary fenestrae absent or extremely small. Maxillopalatine fenestrae and posterolateral foramina present, latter approximately equal to half the width of the last upper molar. Projection of premaxillaries anterior to the incisors is lacking.
Dental characteristics: I1 is longest and separated from I2 by a space. Incisor length increases from I2 through to I5. P2 is larger than P3 and M3 is the widest upper molar. Canines are long and curved.
Skeletal characteristics: Caudal vertebrae vary in number from 32-35.
Genetic characteristics: 2n = 14 Karyotype morphologically similar to the ancestral marsupial complement and can be derived by as few as four changes in centromere position. The nucleolar organizing region (NOR) is located on two pairs of autosomes.
Parasites: Linardi (2006) gives the following ectoparasites recorded in this genus from Brazil: Siphanaptera: Adoratopsylla (Ctenophthalmidae). Acari: Mesostigmata Bdellonyssus (Macronyssidae). Acari: Astigmata Didelphoecius (Atopomelidae).
Paraguayan Species:
Marmosa (Micoureus) constantiae - White-bellied Woolly Mouse Opossum
Marmosa (Micoureus) paraguayana - Long-furred Woolly Mouse Opossum
Tribe Monodelphini Hershkovitz, 1992
Monodelphis Burnett, 1830: Short-tailed Opossums
Twenty-nine species, three present in Paraguay.
Synonyms:
Sorex Müller 1776:36. In part. Not Sorex Linnaeus (1758).
Didelphis Erxleben 1777:80. In part. Not Didelphis Linnaeus (1758).
Didelphys Schreber 1777:549. In part. Unjustified emendation.
Viverra G.Shaw 1800:432. In part. Not Viverra Linnaeus (1758).
Mustela Daudin in Lacépède 1802:163. In part. Not Mustela Linnaeus (1758).
Philander Tiedemann 1808:428. In part.
Sarigua Muirhead 1819:429. In part.
Monodelphis Burnett 1830:351. Type species Monodelphis Brachyura Burnett (1830) (=Didelphys brachyuros Schreber 1777 [=Didelphis brevicaudata Erxleben 1777]) by selection.
? Crossopus Lesson 1842:91. In part. Not Crossopus Wagler (1832).
Micoureus Lesson 1842:186. In part.
Peramys Lesson 1842:187. Type species Peramys brachyurus Lesson (1842) (=Didelphis dimidiata JA Wagner (1847) by selection. Not Didelphis brevicaudata Erxleben (1777).
Grymaeomys Burmeister 1854:138. In part.
Hemiurus P.Gervais 1855:287. Preoccupied.
Microdelphys Burmeister 1856:83. Type species Didelphis (Microdelphys) tristriata Burmeister (1856) (=Didelphys tristriata Illiger 1815, [=Sorex americanus Müller 1776]) by designation. Proposed as a subgenus of Didelphis Linnaeus (1758).
Microdidelphys Trouessart 1898:1242. Incorrect spelling.
Monodelphiops Matschie 1916:261. Type species Microdelphys sorex Hensel (1872) by designation.
Minuania Cabrera 1919:30. Type species Didelphis dimidiata JA Wagner (1847) by description.
Microdelphis Pohle 1927:240. Incorrect spelling.
Monodelphys Reig 1959:57. Incorrect spelling.
Monodelhpis Tálice, Lafitte de Mosera & Machado 1960:151. Incorrect spelling.
General characteristics: Three species of small, shrew-like opossums, with very short tails - 50-70% of the head and body length. Species are extremely similar and distinguished with care, all having short, stout legs, short ears, long-pointed snouts continuous with a steep forehead and a short prehensile-tipped tail. Toes are short, the fifth digit of the hind foot being particularly so and not opposable with the rest of the foot. Claws are long and some species show modifications of the forefeet for digging. Foot pads are poorly developed. Pelage is never long and lax, but varies from short and smooth to long and woolly and guard hairs are absent. Predominately terrestrial and crepuscular in habits, the life cycle is short and neither sex survives to reproduce a second season. No fossil remains have been found that can be assigned to the Pleistocene or earlier.
Cranial characteristics: Palate lacks a second posterior pair of vacuities. Posterolateral foramina reduced. Poorly developed orbitae and lachrimal foramina opening outside the orbit. Alisphenoid with lateral bony rib. Paraoccipital processes well-developed.
Dental characteristics: First premolars not greatly reduced. Third premolar larger or much larger than second premolar on upper and lower jaws. Premolars typically increase in size from front to back. Canine fossae well-developed. Upper first incisor smaller than I2-I4. M4 with vestigial metacone. Lower third premolar larger than p2. m1-4 with short talonids. Poorly developed hypoconids and entoconids. m4 with well-developed cingulum labial to its talonid.
Genetic characteristics: 2n=18. Karyotype can be readily derived from the ancestral marsupial 2n = 14 complement by two fissions and several minor changes in centromere position. NORs have been reported from the short arms of chromosome 5 and the X.
Parasites: This genus shares many ectoparasites with rodents which constitute a major part of the diet. Linardi (2006) gives the following ectoparasites recorded in this genus from Brazil: Siphanaptera: Craneopsylla (Stephanocircidae); Polygenis (Rhopalopsyllidae); Adoratopsylla (Ctenophthalmidae). Acari: Metastigmata Ixodes, Ornithodoros and Amblyomma (Ixodidae). Acari: Mesostigmata Ornithonyssus and Lepronyssoides (Macronyssidae); Eubrachylaelaps, Laelaps Mysolaelaps and Gigantolaelaps (Laelapidae). Acari: Prostigmata Parasecia and Trombewingi (Trombiculidae). Thatcher (2006) documents the following endoparasite for Brazil: Cestoda Linstowia (Anoplocephalidae).
Paraguayan Species:
Monodelphis domestica - Grey Short-tailed Opossum
Monodelphis kunsi - Pygmy Short-tailed Opossum
Monodelphis dimidiata - Red-sided Short-tailed Opossum
Two further species of Monodelphis have been stated to occur in Paraguay in the published literature. M.scalops (Contreras & Silveira 1995) was shown to be an incorrectly identified specimen of M.sorex (de la Sancha et al 2007). Monodelphis brevicaudis mapped for the Paraguayan Chaco by Brown (2004) has variously been regarded to be a senior synonym of M.domestica or M.brevicaudata, but is now generally considered a synonym of M.sorex (de la Sancha et al 2007). Other species may occur in Paraguay but their presence is yet to be proven.
Key to Adults of the Paraguayan Species of Opossum
The fifteen species documented as occurring in Paraguay are keyed as well as three further species considered of hypothetical occurrence: Philander opossum, Micoureus constantiae and Gracilinanus microtarsus.
1a Pelage long and woolly, long prehensile tail furred along basal third, conspicuous medial facial stripe, large forward-facing eyes and arboreal behaviour.................................................................Caluromys lanatus
1b Not as above.....................................................................................................................................................2
2a Size small to medium, terrestrial and typically diurnal in behaviour. Aspect shrew-like or mouse-like with short tail <70% of head and body length........................................................................................... Monodelphis 3
2b Not as above....................................................................................................................................................5
3a Colouration uniform on dorsum without contrasting colouration on flanks, head or rump............................. 4
3b Flanks reddish, contrasting with browner dorsum and paler underparts. Rump dark reddish. Fur short and smooth, lying flat against body....................................................................................Monodelphis dimidiata
4a Size small (hindfoot with claw <16mm). Dorsal colouration uniformly reddish or reddish-brown..........................................................................................................................Monodelphis kunsi
4b Size larger (hindfoot with claw >18mm). Dorsal colouration uniformly grey or grey-brown.....................................................................................................................Monodelphis domestica
5a Conspciuous pale spots above eye and at base of ears..................................................................................... 6
5b No conspicuous pale spots above eye and at base of ears................................................................................8
6a Dorsal pelage predominately brown. Secondary pale spots large and located behind ears. Tail naked at base and uniformly-coloured or paling gradually towards tip - not sharply bicoloured. Female lacks pouch.................................................... ........................................................... Metachirus nudicaudatus
6b Dorsal pelage short and predominately greyish or blackish. Secondary pale spots small and located in front of ears. Tail furred for basal 6-8cm and sharply bicoloured. Pouch present in female................................Philander 7
7a Incomplete labial cingulum on P3. Atlantic Forest............................................................................................................................Philander quica
7b Complete labial cingulum on P3. Chaco and Paraguay River Basin. ...........................................................................................................................................Philander canus
8a Dorsal pelage dense and velvety, predominately black with grey markings. Tail furred for basal 10%, naked and black for rest of length with whitish tip. Webbed feet. Swims well......................Chironectes minimus
8b Not as above....................................................................................................................................................9
9a Ears extremely short, rounded and dark, barely projecting above fur. General appearance weasel-like. Pelage uniformly brownish or reddish. Tail extremely thick at base, and furred along basal third. Mainly terrestrial but climbs and swims readily.....................................................................................Lutreolina crassicaudata
9b Not as above...................................................................................................................................................10
10a Size large (total length >500mm). Pelage with long, prominent guard hairs and soft, dense underfur. Females with well-developed marsupium..............................................................................................Didelphis 11
10b Size small or medium (total length generally much less than 450mm) . General appearance mouse-like. Pouch absent........................................................................................................................................................12
11a Dorsal pelage mostly greyish. Prominent white facial markings. Ears pale. Widespread in a variety of habitats.......................................................................................................................Didelphis albiventris
11b Dorsal pelage mostly blackish. Golden-buff facial markings. Ears dark. Forested areas only................................................................................................................................Didelphis aurita
12a Size medium (TL>350mm), white tip to tail..................................................................................Micoureus 13
12b Size small (TL <200mm). Pelage short. Usually no white tip to tail.............................................................14
13a Fur long and woolly extending 30mm or so down base of tail. Ventral pelage buffy to cinnamon and grey-based. Southern and eastern Paraguay in Atlantic Forest....................................Micoureus paraguayanus
13b Fur long but not overly dense and not notably extending onto base of tail. Ventral pelage creamy to yellow-white and white-based. In Chaco and Cerrado.....................Micoureus constantiae
14a Ratio of tail length to head and body length between 1 and 1.25. Fat may be stored at base of tail at certain times of year. Soles of feet densely covered with small tubercles. Length of digit IV <0.45 times the hindfoot length. Teats arranged in bilaterally symmetrical rows...........................................................................Thylamys 15
14b Ratio of tail length to head and body length between 1 and 1.96. Fat never stored at base of tail. Tail scales rounded or square with annular arrangement. Soles of feet smooth or sparsely granulated. Length of digit IV >0.45 times the hindfoot length. Teats with typical circular arrangement..............................................................16
15a Size large, tail >135mm. Weight >40g. Tail with whitish tip. Orient only...........Thylamys macrurus
15b Size small, tail <135mm. Weight <35g. Tail lacks whitish tip. Chaco only...........Thylamys pusillus
16a Ventral pelage creamy with greyish base. Ratio of tail length to head and body length >1.2 but usually <1.5. Tail length 110-165mm. P2 and P3 of equal height..........................................................................Gracilinanus 17
16b Ventral pelage not grey-based. Ratio of tail length to head and body length <1.2. Tail length 95-117mm. P2 smaller than P3......................................................................................................Cryptonanus chacoensis / unduaviensis.
17a Dorsal pelage pale brown to greyish-brown looking grizzled. Ventral pelage grey-based except for chin, throat, upper breast and scrotal area. Ocular patch does not extend to nose and ears. Face not markedly paler than dorsum. Tail typically <140mm. Ears usually >21mm. Mainly cerrado in distribution..................................................................................................................Gracilinanus agilis
17b Dorsal pelage reddish-brown to chestnut-brown looking uniform. Ventral entirely grey-based except for chin. Ocular patch extends to nose and ears. Face contrastingly paler than dorsum. Tail typically >140mm. Ears usually <21mm. Hypothetical in Atlantic Forest..................................................................Gracilinanus microtarsus
REFERENCES
Brown BE 2004 - Atlas of New World Marsupials - Fieldiana Zoology 1257.






















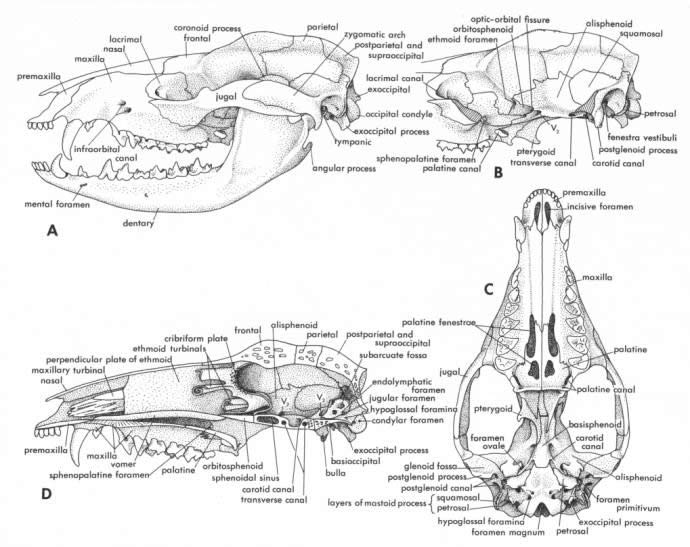
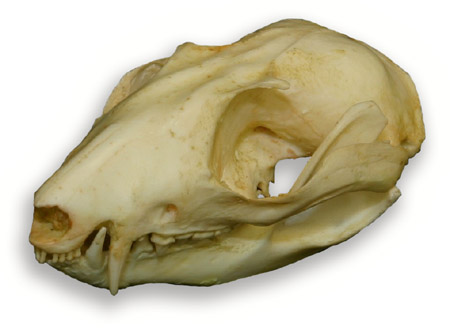
FIG 1: Skull Caluromys lanatus. Click the image to enlarge it.
Image courtesy of www.skullsunlimited.com
Image courtesy of www.skullsunlimited.com
Click on the species links for access to the image gallery.
Click on the HBook icon on the right hand side for access to pdfs of the FAUNA Paraguay Handbook of the Mammals of Paraguay Volume 1 Didelphimorphia.
See below for a key to the Paraguayan species.
MAGNORDER AMERIDELPHIA - NEW WORLD METATHERIANS
ORDER DIDELPHIMORPHIA - DIDELPHIDS
FAMILY DIDELPHIDAE - OPOSSUMS
General characteristics: Sixteen species in one family. There is much variation in size amongst the genera, from very small (Cryptonanus 12cm; 15g) to large (Didelphis 105cm; 2kg). The snout is long and in most species the ears are well-developed though delicate, often being smoothed down against the head whilst walking to avoid damage. Terrestrial opossums are characterized by a comparatively short tail and well-developed hind legs, arboreal species by a long tail (often with a prehensile tip) and large forelimbs. Each foot possesses five digits, the forefeet small, the hind feet slightly longer with a large opposable thumb lacking a nail.
Cranial characteristics: Robust zygomatic arch. Elongated cranial case. Angular process of mandible bent inwards. Small auditory bullae are tripartite consisting of unfused elements of the alisphenoid, periotic and ectotympanic. A fenestrated bony palate is diagnostic of the family. See Fig 3 for annotated Didelphid skull.
Dental characteristics: Dental formula I5/4 C1/1 P3/3 M4/4 = 50. The dentition is heterodont with as many as 50 teeth typical, all teeth behind the canines being pointed. Incisors polyprotodont, I1 being longest and separated from 12 by a space. Upper canines well-developed. Third premolar small multicuspid, molariform and deciduous. Molars trituberculate with prominent stylar shelf. Ageing of specimens is most effective when dental classes are based on the unvarying linear sequence of upper molar eruption, as individual variation in eruption sequence of other teeth is not uncommon, even among members of the same litter (Van Nievelt & Smith 2006).
Skeletal characteristics: Pentadactyl feet with a phalangeal formula 2-3-3-3-3. Hallux lacks a nail but retains a reduced eponychium.
Genetic characteristics: 2n=22 in Didelphis, Lutreolina, Chironectes and Philander; 2n=18 in Monodelphis; 2n=14 in Cryptonanus, Gracilinanus, Metachirus, Micoureus and Thylamys.
Ecology: Though rarely observed, Didelphids are often amongst the most numerous of South American mammals in any given community. Most species are nocturnal (Monodelphis is a notable exception) and can be detected by the bright red or orange eye-shine they give under torchlight, the eyes typically appearing small and well-separated. Didelphids are typically omnivorous, eating a mixture of plant and animal matter, though the relative proportions of each in the diet varies considerably between species. Vieira & Astúa de Morães (2003) classified the Ameridelphia into 5 diet classes, with class 1 representing the most frugivorous and class 5 representing the most carnivorous with the following results: Class 1 Caluromys; 2 Micoureus, Gracilinanus, Cryptonanus; 3 Didelphis; 4 Philander, Metachirus, Monodelphis, Thylamys; 5 Lutreolina, Chironectes. Several species with apparently similar ecology are able to co-exist in forested habitats by varying the proportions of foodstuffs in their diet and by occupying different levels of the forest canopy; some species being exclusively terrestrial, other exclusively arboreal and others occupying different microhabitats between the two. The role of some Didelphids in seed dispersal of forest plants has recently become apparent. See Cáceres (2006) for an excellent review of processes involved.
Reproductive Biology: The reproductive behaviour of this group is unique amongst Paraguayan mammals, the young being born undeveloped but with well-developed forelimbs after a brief gestation of a couple of weeks. The newborn then makes its way to the teat where it continues its development for a further 4 to 6 weeks. Once they reach a size too large to be transported by the mother they are deposited in a nest lined with dry leaves where they continue to be fed. A pouch is present in some genera (Didelphis, Philander, Chironectes etc) but absent in others (Monodelphis, Metachirus, mouse-opossums), while Caluromys develops a pouch only during the period of lactation. Nogueira (2003) and Nogueira & Castro (2003) describe the male genital morphology of opossums and Montero Filho & Cáceres (2003) provide a rundown of female reproductive biology.
Physiology: See Quadros (2003) for a microscopic analysis of guard hairs of Brazilian Didelphimorphia and its utility for species identification. José (2003) gives a description of paracloacal glands. Hokoç et al (2006) review the characteristics and vision of Brazilian species.
Parasites: Genera and species of parasites documented in species occurring in Paraguay are given in the generic and species accounts.
Evolution of South American "Marsupials": (adapted from Tyndale-Briscoe 2005): South America´s isolation from North America at the end of the Cretaceous 65 million years ago left the continent´s mammals to evolve independently for the next 35 million years. With placental mammals assuming the herbivorous role, the marsupials filled the ecological niche of small insectivore and shared the niche of large carnivore with birds. An almost complete fossil history allows us to trace the lineages of the present day marsupials through the whole span of the Tertiary.
In one extraordinarily rich fossil site at Tiupampa, Bolivia considered to be very early Tertiary, 11 species of marsupials and seven species of placental are known from teeth, skulls and even whole skeletons, demonstrating that the lineages that would predominate for the next 65 million years were already established in early history. In the Palaeocene and Eocene epochs, which encompass the first 25 million years of the Tertiary, 24 genera of marsupials representing six distinct families are known. The one herbivorous group, the polydolopodids, did not survive past the Eocene but the carnivorous species persisted.
Total South American isolation occurred in the Oligocene (30 million years ago) when the continent was invaded by the earliest rodents and primates. They may have displaced some of the smaller arboreal marsupials or even prevented marsupials from evolving into the small to medium herbivore niches that they came to dominate thereafter. At round this time in Australia, a country with a dominant marsupial fauna, a great expansion of marsupials into arboreal and browsing niches took place.
In the Miocene (23-25 million years ago) the caenolestids (which survive today) and the large carnivorous borhyaenids, became abundant. The borhyaenids flourished until the Pleistocene epoch (3 million years ago). Borhyaenids were the large mammalian carnivores of the continent for more than 20 million years. Their short limbs made them stocky creatures like badgers, not lithe pursuit hunters like wolves. A bizarre case of convergent evolution was the sabre-tooth borhyaenids that flourished in the Pliocene (5 million years ago), at the same time as placental sabre-tooth tigers, Barbourofelis and Smilodon, were living in North America. Thylacosmilus atrox was a large carnivore with a pair of huge upper canines with very thin enamel and open roots, so that they could grow continuously through life. Its teeth and skull were similar to those of the northern sabre-tooth tigers but its short, strong forelimbs were suggestive of an ambush hunter that stabbed its prey to death, helped by its enormous (67°) gape. Both the variety of prey species and the marsupials that hunted them disappeared just before or soon after the continent became joined to North America about 3 million years ago.
In the Pliocene epoch North and South America rejoined for the first time in more than 60 million years. Once the land connection was established a flood of species moved south and a lesser number of species moved north: a phenomenon known as the great American faunal interchange. It is not clear to what extent the invasion affected the indigenous marsupials but all the large carnivorous marsupials became extinct at or before this time. The shrews and moles (Order Insectivora) were one of the few groups that failed to penetrate the southern continent, perhaps because the small insectivore niche was already fully occupied by small to medium marsupials. Small placental carnivores, ancestors of the Procyonidae, deeply penetrated into South America however, and possibly contributed to the decline and extinction of the borhyaenids. Around the same time, the didelphids, began to flourish and present day genera began to appear. Smaller species, such as Monodelphis first appeared in the Miocene, Lutreolina, Thylamys and Philander somewhat later in the Pliocene and other species, including Didelphis appear in the Pleistocene, just 2 million years ago. A parallel evolution occurred in Australia, where the early carnivores were large thylacinids that were replaced in the last five million years by the smaller dasyurids.
Another family of marsupials appeared in the Pliocene: Argyrolagus were small, hopping forms, with very short forelegs and elongated hind. They were similar to modern day hopping mice but became extinct in the Pleistocene about 2 million years ago.
The fossil history suggests that the present day marsupial families are not closely related, this being corroborated by recent molecular evidence. On these criteria the Microbiotheriidae, represented today by Dromiciops, and the Didelphidae separated more than 60 million years ago, during the Palaeocene. Likewise, the differences between the Caenolestidae and Didelphidae put their separation at about 50 million years ago.
For more information on early evolution of Marsupials in South America see Goin (2003).
Taxonomy: Metatherians (more commonly known as marsupials) where until recently classified in the single order Marsupialia, which included both Australasian and Neotropical species. More recently molecular work has revealed that in fact the group is made up of seven separate orders, three of which are Neotropical (Didelphimorphia, Paucituberculata and Microbiotheria) and one of which occurs in Paraguay. Higher level taxonomy of Neotropical Didelphimorphia is much disputed. According to Reig, Kirsch & Marshall (1985) the single family Didelphidae could be separated into two subfamilies - Caluromyinae and Didelphinae. The former represented in Paraguay only by a single species in the genus Caluromys, the latter split into three tribes Metachirini (containing only Metachirus), Monodelphini (containing the genera Monodelphis, Cryptonanus, Gracilinanus, Micoureus and Thylamys) and Didelphini (containing the genera Chironectes, Didelphis, Lutreolina and Philander). This classification was followed by Gardner (2007). Hershkovitz (1992) raised Caluromyidae (including Caluromys) and Marmosidae (including Monodelphis, Cryptonanus, Gracilinanus, Metachirus, Micoureus and Thylamys) to family level, though this arrangement failed to gain much support and worked on the assumption that all small "mouse opossums" were closely related. More recently DNA-DNA hybridisation studies have recognised Caluromyidae as a separate family and split Didelphidae into three subfamilies: Didelphinae (containing the genera Chironectes, Didelphis, Lutreolina, Metachirus and Philander), Marmosinae ("pouchless mouse opossums" containing the genera Micoureus and Monodelphis) and Thylamyinae ("pouched mouse opossums" containing the genera Cryptonanus, Gracilinanus and Thylamys) (Kirsch & Palma 1995). The arrangement here follows the results of molecular work by Jansa et al (2006) and Jansa & Voss (2000).
Web Resources: The following online resources are available:
Specimen data file for Didelphids collected at Reserva Bosque Mbaracayú.
Website of Professor Rui Cerqueira - Has published widely on Brazilian Didelphidae with pdfs to download.
Subfamily Caluromyinae Kirsch, 1977
Cranial characteristics: Palatal fenestrae and vacuities absent.
Dental characteristics: M2 longer than or of equal length to M3. Upper molars separated with subequal paracones and metacones and paracrista not united to stylar cusps.
Taxonomy: Five species in three genera in South America. A single species in the genus Caluromys occurs in Paraguay. This subfamily is sometimes raised to family level.
Caluromys JA Allen, 1900: Woolly Opossums
Three species one in Paraguay. Synonyms adapted from Gardner (2007).
Synonyms:
Didelphis Linnaeus 1758:54. In part.
Philander Beckmann 1772:244. Type species Didelphis philander Linnaeus 1758 by absolute tautonomy. Preoccupied by Philander Brisson 1762.
Didelphys Schreber 1777:532. In part. Unjustified emendation.
Sarigua Muirhead 1819:429. In part.
Micoureus Lesson 1842:186. In part.
Philander Burmeister 1856:74. Type species Didelphys (Philander) cayopollin, Burmeister 1856 (=Didelphys cayopollin Schreber 1778). Described as a subgenus of Didelphis. Name preoccupied.
Gamba Liais 1872:330. In part.
Cuica Liais 1872:330. In part.
Micoureus Ihering 1894:11. In part.
Caluromys JA Allen 1900:189. Type species Didelphis philander Linnaeus 1758.
Micoureus Matschie 1916:269. In part. Described as a subgenus of Didelphis.
Mallodelphys O.Thomas 1920:195. Type species Didelphis laniger Desmarest 1820. Described as subgenus of Philander.
Calaromys A.Miranda-Ribeiro 1936:324. Incorrect spelling.
Mallodelphis Gilmore 1941:317. Incorrect spelling.
Calurosmys Ávila-Pires 1964:11. Incorrect spelling.
General characteristics: A single, strictly arboreal species in Paraguay with extremely thick, woolly pelage and dark facial stripes. Tail very long and prehensile, furred along the basal one-third of its length. Ears large and naked. Captive specimens live longer than other Didelphids and studies reveal that extended maternal care and small litter sizes are the norm. They have a larger brain volume than other members of the family. Females develop a pouch only for the period of lactation. Fossil material is known from the Pleistocene of South America. Diploid chromosome number 14.
Cranial characteristics: (Fig 1) Skull is short and broad with well-developed supraorbital processes, large orbits, a broad brain case and a broad palate lacking fenestrae.
Dental characteristics: Stylar shelf of the upper molars reduced and stylar cusps reduced.
Genetic characteristics: 2n=14. In Caluromys the nucleolar organizing region (NOR) is located on a single pair of autosomes.
Parasites: Thatcher (2006) documents the Protozoa Trypanosoma for Brazil.
Paraguayan Species:
Caluromys lanatus - Brown-eared Woolly Opossum HBook 16
Subfamily Didelphinae Gray, 1821
Tribe Didelphini Gray, 1821
General characteristics: Five large opossums in four genera with total length >500mm. Pouch moderately to well-developed.
Didelphis Linnaeus, 1758: True Opossums
Five species, two of which occur in Paraguay. Synonyms adapted from Gardner (2007).
Synonyms:
Didelphis Linnaeus 1758:54. Type species Didelphis marsupialis Linnaeus (1758).
Didelphys Schreber 1777:532. Unjustified emendation.
Opossum Schmid 1818:115. In part. Didelphis marsupialis Linnaeus (1758).
Sarigua Muirhead 1819:429. In part.
Didelphus I. Geoffroy St.Hilaire 1831:139. Incorrect spelling.
Thylacotherium Lund 1839:233. Type species Thylacotherium ferox Lund (1839). Preoccupied by Thylacotherium Valenciennes 1838.
Micoureus Lesson 1842:186. In part.
Didelphus Lapham 1853:337. Incorrect spelling.
Gamba Liais 1872:329. In part.
Gambatherium Liais 1872:331. Replacement name for Thylacotherium Lund (1839).
Dasyurotherium Liais 1872:331. Replacement name for Thylacotherium Lund (1839).
Dimerodon Ameghino 1889:277. Type species Dimerodon mutilatus Ameghino (1889) by monotypy.
Leucodidelphis Ihering 1914:347. Type species Didelphis paraguayensis Oken (1816). Name not available. Proposed as subgenus of Didelphis.
Leucodidelphys Krumbiegel 1941:34. Unjustified emendation.
Leucodelphis Cabrera 1958:41. Incorrect spelling.
General characteristics: Two species of "typical" opossum with pointed snout, large, naked, rounded ears and long, prehensile tail furred at base and white towards the tip. This genus includes the largest of New World marsupials. Females have a well-developed pouch with 11 to 13 teats. Pelage is soft and dense with long, coarse guard hairs. Semi-arboreal in behaviour. Fossils from the middle Pleistocene of South America have been referred to this genus
Cranial characteristics: Well-developed sagittal crest in adults. Narrow braincase with interorbital width greater than the postorbital constriction.
Dental characteristics: Dental wear can be used for aging in this genus, see species accounts and (Fig 2).
Genetic characteristics: 2n=14. The nucleolar organizing region (NOR) is located on up to three pairs of autosomes.
Parasites: Linardi (2006) gives the following ectoparasites recorded in this genus from Brazil: Siphanaptera: Craneopsylla (Stephanocircidae); Ctenocephalides (Pulicidae); Polygenis and Rhopalopsyllus (Rhopalopsyllidae); Adoratopsylla (Ctenophthalmidae). Acari: Metastigmata Ixodes, Ornithodoros and Amblyomma (Ixodidae). Acari: Mesostigmata Ornithonyssus and Bdellonyssus (Macronyssidae); Androlaelaps, Laelaps and Gigantolaelaps (Laelapidae). Acari: Prostigmata Archemyobia (Myobiidae). Acari: Astigmata Didelphilichus (Atopomelidae). Thatcher (2006) documents the following endoparasites for Brazil: Protozoa Trypanosoma and Babesia. Trematoda Schistosoma (Schistosomatidae), Aphimerus (Opisthorchiidae), Rhopalias (Rhopaliasidae) and Paragonimus (Paragonimidae). Cestoda Mathevotaenia bivittata (Anoplocephalidae). Nematoda Dracunculus (Dracunculidae); Gongelonemoides (Gongylonematidae); Dipetalonema (Oncocercidae); Thominx (Trichuridae); Travassostrongylus and Viannaia (Trichostrongylidae). Acanthocephala Oligacanthorhynchus (Oligacanthorhynchidae).
Evolution: Formerly considered to be the most primitive of Didelphid lineages, Patton & Costa (2003) demonstrated that the genus is in fact young and speciated relatively recently.
Paraguayan Species:
Didelphis albiventris - White-eared Opossum
Didelphis aurita - Southern Black-eared Opossum
Chironectes Illiger, 1811: Water Opossum
This is a monotypic genus. Synonyms adapted from Gardner (2007).
Synonyms:
Latra Zimmermann 1780:317. In part. Incorrect spelling of Lutra. Not Lutra Linnaeus 1758.
Mustela Kerr 1792:172. In part. Not Mustela Linnaeus 1758.
Lutra Link 1795:172. Not Lutra Linnaeus 1758.
Didelphis G.Cuvier 1798:125. In part. Not Didelphis Linnaeus 1758.
Lutra G.Shaw 1800:447. Not Lutra Linnaeus 1758.
Mustela Turton 1800:58. In part. Not Mustela Linnaeus 1758.
Didelphis Daudin in Lacépède 1802:152. Not Didelphis Linnaeus 1758.
Chironectes Illiger 1811:76. Type species Lutra minima Zimmermann 1780 by monotypy.
Memina G.Fischer 1813:15. Nomen nudum.
Memina G.Fischer 1814:11. Type species Lutra memina Boddaert 1784 by monotypy.
Sarigua Muirhead 1819:429. In part.
Cheironectes Gray 1821:308. Incorrect spelling.
Cheronectis Fleming 1822:212. Incorrect spelling.
Gamba Liais 1872:329. Type species Gamba palmata Liais 1872 by designation.
Chironeytes Goeldi & Hagman 1904:100. Incorrect spelling.
General characteristics: Chironectes is the only truly aquatic opossum. The tail is longer than the head and body, naked and rounded in cross-section. It is not prehensile. The ears are fairly large, rounded and naked. The snout is long with stiff facial bristles. Further adaptations for an aquatic existence include the dense, short, waterproof pelage and the hindfeet webbed to the tips of the toes. The forefeet are unwebbed with long, dextrous fingers, clubbed at the end. A well-developed, backwards-opening pouch is present in both sexes and can be sealed whilst the animal is submerged. There are 4 or 5 mammae. Fossils from the Miocene of Argentina have been referred to this genus.
Cranial characteristics: Recalls that of Philander. Long rostrum, rounded braincase and broadly flared zygomatic arches. Palate with a single pair of maxillopalatine fenestrae opposite the molars. Small palatine fenestrae are absent.
Genetic characteristics: 2n=14. The nucleolar organizing region (NOR) is located on a single pair of autosomes.
Parasites: Linardi (2006) gives the following ectoparasites recorded in this genus from Brazil: Siphanaptera: Adoratopsylla (Ctenophthalmidae). Thatcher (2006) documents the following endoparasites for Brazil: Trematoda Rhopalias (Rhopaliasidae).
Paraguayan Species:
Chironectes minimus - Water Opossum
Lutreolina O. Thomas, 1910: Thick-tailed Opossum
This is a monotypic genus. Synonyms adapted from Gardner (2007).
Synonyms:
Didelphis Desmarest 1804:19. In part. Not Didelphis Linnaeus (1758).
Didelphys Olfers 1818:204. In part. Incorrect spelling.
Sarigua Muirhead 1819:429. In part.
Peramys Lesson 1842:261. In part.
Micoureus P.Gervais 1855:287. In part. Not Micoureus Lesson (1842).
Philander Gerard 1862:139. In part. Not Philander Brisson (1762).
Metachirus Hensel 1872:121. In part. Proposed as a subgenus of Didelphis. Not Metachirus Burmeister (1854).
Lutreolina O.Thomas 1910:247. Type species Didelphis crassicaudata Desmarest (1804) by monotypy.
General characteristics: Lutreolina bears a superficial resemblance to a small otter or weasel, with a thick, non-prehensile tail furred to approximately half its length. Pelage is uniformly-coloured, thick and dense but not waterproof. Living or freshly-killed specimens fluoresce red-orange under UV light. The snout is short and the ears small and rounded. The legs are short and strong. This species possesses only a slight pouch. Hallux is not fully opposable. Fossils from the Pliocene of Argentina have been referred to this genus
Cranial characteristics: Shows several adaptations related to a more carnivorous diet. Distinctive amongst Didelphids on account of the shortened rostrum, narrow braincase and zygomatic arches. The nasals taper evenly. Braincase narrower and longer than similar-sized species with widened temporal fossa and broader, more perpendicular coronoid processes. (Vieira & Astúa de Morães 2003)
Genetic characteristics: 2n=14. The nucleolar organizing region (NOR) is located on two pairs of autosomes.
Parasites: Linardi (2006) gives the following ectoparasites recorded in this genus from Brazil: Siphanaptera: Craneopsylla (Stephanocircidae); Polygenis (Rhopalopsyllidae); Ctenocephalides (Pulicidae); Adoratopsylla (Ctenophthalmidae). Acari: Metastigmata Ixodes (Ixodidae).
Paraguayan Species:
Lutreolina crassicaudata - Thick-tailed Opossum
Click on the HBook icon on the right hand side for access to pdfs of the FAUNA Paraguay Handbook of the Mammals of Paraguay Volume 1 Didelphimorphia.
See below for a key to the Paraguayan species.
MAGNORDER AMERIDELPHIA - NEW WORLD METATHERIANS
ORDER DIDELPHIMORPHIA - DIDELPHIDS
FAMILY DIDELPHIDAE - OPOSSUMS
General characteristics: Sixteen species in one family. There is much variation in size amongst the genera, from very small (Cryptonanus 12cm; 15g) to large (Didelphis 105cm; 2kg). The snout is long and in most species the ears are well-developed though delicate, often being smoothed down against the head whilst walking to avoid damage. Terrestrial opossums are characterized by a comparatively short tail and well-developed hind legs, arboreal species by a long tail (often with a prehensile tip) and large forelimbs. Each foot possesses five digits, the forefeet small, the hind feet slightly longer with a large opposable thumb lacking a nail.
Cranial characteristics: Robust zygomatic arch. Elongated cranial case. Angular process of mandible bent inwards. Small auditory bullae are tripartite consisting of unfused elements of the alisphenoid, periotic and ectotympanic. A fenestrated bony palate is diagnostic of the family. See Fig 3 for annotated Didelphid skull.
Dental characteristics: Dental formula I5/4 C1/1 P3/3 M4/4 = 50. The dentition is heterodont with as many as 50 teeth typical, all teeth behind the canines being pointed. Incisors polyprotodont, I1 being longest and separated from 12 by a space. Upper canines well-developed. Third premolar small multicuspid, molariform and deciduous. Molars trituberculate with prominent stylar shelf. Ageing of specimens is most effective when dental classes are based on the unvarying linear sequence of upper molar eruption, as individual variation in eruption sequence of other teeth is not uncommon, even among members of the same litter (Van Nievelt & Smith 2006).
Skeletal characteristics: Pentadactyl feet with a phalangeal formula 2-3-3-3-3. Hallux lacks a nail but retains a reduced eponychium.
Genetic characteristics: 2n=22 in Didelphis, Lutreolina, Chironectes and Philander; 2n=18 in Monodelphis; 2n=14 in Cryptonanus, Gracilinanus, Metachirus, Micoureus and Thylamys.
Ecology: Though rarely observed, Didelphids are often amongst the most numerous of South American mammals in any given community. Most species are nocturnal (Monodelphis is a notable exception) and can be detected by the bright red or orange eye-shine they give under torchlight, the eyes typically appearing small and well-separated. Didelphids are typically omnivorous, eating a mixture of plant and animal matter, though the relative proportions of each in the diet varies considerably between species. Vieira & Astúa de Morães (2003) classified the Ameridelphia into 5 diet classes, with class 1 representing the most frugivorous and class 5 representing the most carnivorous with the following results: Class 1 Caluromys; 2 Micoureus, Gracilinanus, Cryptonanus; 3 Didelphis; 4 Philander, Metachirus, Monodelphis, Thylamys; 5 Lutreolina, Chironectes. Several species with apparently similar ecology are able to co-exist in forested habitats by varying the proportions of foodstuffs in their diet and by occupying different levels of the forest canopy; some species being exclusively terrestrial, other exclusively arboreal and others occupying different microhabitats between the two. The role of some Didelphids in seed dispersal of forest plants has recently become apparent. See Cáceres (2006) for an excellent review of processes involved.
Reproductive Biology: The reproductive behaviour of this group is unique amongst Paraguayan mammals, the young being born undeveloped but with well-developed forelimbs after a brief gestation of a couple of weeks. The newborn then makes its way to the teat where it continues its development for a further 4 to 6 weeks. Once they reach a size too large to be transported by the mother they are deposited in a nest lined with dry leaves where they continue to be fed. A pouch is present in some genera (Didelphis, Philander, Chironectes etc) but absent in others (Monodelphis, Metachirus, mouse-opossums), while Caluromys develops a pouch only during the period of lactation. Nogueira (2003) and Nogueira & Castro (2003) describe the male genital morphology of opossums and Montero Filho & Cáceres (2003) provide a rundown of female reproductive biology.
Physiology: See Quadros (2003) for a microscopic analysis of guard hairs of Brazilian Didelphimorphia and its utility for species identification. José (2003) gives a description of paracloacal glands. Hokoç et al (2006) review the characteristics and vision of Brazilian species.
Parasites: Genera and species of parasites documented in species occurring in Paraguay are given in the generic and species accounts.
Evolution of South American "Marsupials": (adapted from Tyndale-Briscoe 2005): South America´s isolation from North America at the end of the Cretaceous 65 million years ago left the continent´s mammals to evolve independently for the next 35 million years. With placental mammals assuming the herbivorous role, the marsupials filled the ecological niche of small insectivore and shared the niche of large carnivore with birds. An almost complete fossil history allows us to trace the lineages of the present day marsupials through the whole span of the Tertiary.
In one extraordinarily rich fossil site at Tiupampa, Bolivia considered to be very early Tertiary, 11 species of marsupials and seven species of placental are known from teeth, skulls and even whole skeletons, demonstrating that the lineages that would predominate for the next 65 million years were already established in early history. In the Palaeocene and Eocene epochs, which encompass the first 25 million years of the Tertiary, 24 genera of marsupials representing six distinct families are known. The one herbivorous group, the polydolopodids, did not survive past the Eocene but the carnivorous species persisted.
Total South American isolation occurred in the Oligocene (30 million years ago) when the continent was invaded by the earliest rodents and primates. They may have displaced some of the smaller arboreal marsupials or even prevented marsupials from evolving into the small to medium herbivore niches that they came to dominate thereafter. At round this time in Australia, a country with a dominant marsupial fauna, a great expansion of marsupials into arboreal and browsing niches took place.
In the Miocene (23-25 million years ago) the caenolestids (which survive today) and the large carnivorous borhyaenids, became abundant. The borhyaenids flourished until the Pleistocene epoch (3 million years ago). Borhyaenids were the large mammalian carnivores of the continent for more than 20 million years. Their short limbs made them stocky creatures like badgers, not lithe pursuit hunters like wolves. A bizarre case of convergent evolution was the sabre-tooth borhyaenids that flourished in the Pliocene (5 million years ago), at the same time as placental sabre-tooth tigers, Barbourofelis and Smilodon, were living in North America. Thylacosmilus atrox was a large carnivore with a pair of huge upper canines with very thin enamel and open roots, so that they could grow continuously through life. Its teeth and skull were similar to those of the northern sabre-tooth tigers but its short, strong forelimbs were suggestive of an ambush hunter that stabbed its prey to death, helped by its enormous (67°) gape. Both the variety of prey species and the marsupials that hunted them disappeared just before or soon after the continent became joined to North America about 3 million years ago.
In the Pliocene epoch North and South America rejoined for the first time in more than 60 million years. Once the land connection was established a flood of species moved south and a lesser number of species moved north: a phenomenon known as the great American faunal interchange. It is not clear to what extent the invasion affected the indigenous marsupials but all the large carnivorous marsupials became extinct at or before this time. The shrews and moles (Order Insectivora) were one of the few groups that failed to penetrate the southern continent, perhaps because the small insectivore niche was already fully occupied by small to medium marsupials. Small placental carnivores, ancestors of the Procyonidae, deeply penetrated into South America however, and possibly contributed to the decline and extinction of the borhyaenids. Around the same time, the didelphids, began to flourish and present day genera began to appear. Smaller species, such as Monodelphis first appeared in the Miocene, Lutreolina, Thylamys and Philander somewhat later in the Pliocene and other species, including Didelphis appear in the Pleistocene, just 2 million years ago. A parallel evolution occurred in Australia, where the early carnivores were large thylacinids that were replaced in the last five million years by the smaller dasyurids.
Another family of marsupials appeared in the Pliocene: Argyrolagus were small, hopping forms, with very short forelegs and elongated hind. They were similar to modern day hopping mice but became extinct in the Pleistocene about 2 million years ago.
The fossil history suggests that the present day marsupial families are not closely related, this being corroborated by recent molecular evidence. On these criteria the Microbiotheriidae, represented today by Dromiciops, and the Didelphidae separated more than 60 million years ago, during the Palaeocene. Likewise, the differences between the Caenolestidae and Didelphidae put their separation at about 50 million years ago.
For more information on early evolution of Marsupials in South America see Goin (2003).
Taxonomy: Metatherians (more commonly known as marsupials) where until recently classified in the single order Marsupialia, which included both Australasian and Neotropical species. More recently molecular work has revealed that in fact the group is made up of seven separate orders, three of which are Neotropical (Didelphimorphia, Paucituberculata and Microbiotheria) and one of which occurs in Paraguay. Higher level taxonomy of Neotropical Didelphimorphia is much disputed. According to Reig, Kirsch & Marshall (1985) the single family Didelphidae could be separated into two subfamilies - Caluromyinae and Didelphinae. The former represented in Paraguay only by a single species in the genus Caluromys, the latter split into three tribes Metachirini (containing only Metachirus), Monodelphini (containing the genera Monodelphis, Cryptonanus, Gracilinanus, Micoureus and Thylamys) and Didelphini (containing the genera Chironectes, Didelphis, Lutreolina and Philander). This classification was followed by Gardner (2007). Hershkovitz (1992) raised Caluromyidae (including Caluromys) and Marmosidae (including Monodelphis, Cryptonanus, Gracilinanus, Metachirus, Micoureus and Thylamys) to family level, though this arrangement failed to gain much support and worked on the assumption that all small "mouse opossums" were closely related. More recently DNA-DNA hybridisation studies have recognised Caluromyidae as a separate family and split Didelphidae into three subfamilies: Didelphinae (containing the genera Chironectes, Didelphis, Lutreolina, Metachirus and Philander), Marmosinae ("pouchless mouse opossums" containing the genera Micoureus and Monodelphis) and Thylamyinae ("pouched mouse opossums" containing the genera Cryptonanus, Gracilinanus and Thylamys) (Kirsch & Palma 1995). The arrangement here follows the results of molecular work by Jansa et al (2006) and Jansa & Voss (2000).
Web Resources: The following online resources are available:
Specimen data file for Didelphids collected at Reserva Bosque Mbaracayú.
Website of Professor Rui Cerqueira - Has published widely on Brazilian Didelphidae with pdfs to download.
Subfamily Caluromyinae Kirsch, 1977
Cranial characteristics: Palatal fenestrae and vacuities absent.
Dental characteristics: M2 longer than or of equal length to M3. Upper molars separated with subequal paracones and metacones and paracrista not united to stylar cusps.
Taxonomy: Five species in three genera in South America. A single species in the genus Caluromys occurs in Paraguay. This subfamily is sometimes raised to family level.
Caluromys JA Allen, 1900: Woolly Opossums
Three species one in Paraguay. Synonyms adapted from Gardner (2007).
Synonyms:
Didelphis Linnaeus 1758:54. In part.
Philander Beckmann 1772:244. Type species Didelphis philander Linnaeus 1758 by absolute tautonomy. Preoccupied by Philander Brisson 1762.
Didelphys Schreber 1777:532. In part. Unjustified emendation.
Sarigua Muirhead 1819:429. In part.
Micoureus Lesson 1842:186. In part.
Philander Burmeister 1856:74. Type species Didelphys (Philander) cayopollin, Burmeister 1856 (=Didelphys cayopollin Schreber 1778). Described as a subgenus of Didelphis. Name preoccupied.
Gamba Liais 1872:330. In part.
Cuica Liais 1872:330. In part.
Micoureus Ihering 1894:11. In part.
Caluromys JA Allen 1900:189. Type species Didelphis philander Linnaeus 1758.
Micoureus Matschie 1916:269. In part. Described as a subgenus of Didelphis.
Mallodelphys O.Thomas 1920:195. Type species Didelphis laniger Desmarest 1820. Described as subgenus of Philander.
Calaromys A.Miranda-Ribeiro 1936:324. Incorrect spelling.
Mallodelphis Gilmore 1941:317. Incorrect spelling.
Calurosmys Ávila-Pires 1964:11. Incorrect spelling.
General characteristics: A single, strictly arboreal species in Paraguay with extremely thick, woolly pelage and dark facial stripes. Tail very long and prehensile, furred along the basal one-third of its length. Ears large and naked. Captive specimens live longer than other Didelphids and studies reveal that extended maternal care and small litter sizes are the norm. They have a larger brain volume than other members of the family. Females develop a pouch only for the period of lactation. Fossil material is known from the Pleistocene of South America. Diploid chromosome number 14.
Cranial characteristics: (Fig 1) Skull is short and broad with well-developed supraorbital processes, large orbits, a broad brain case and a broad palate lacking fenestrae.
Dental characteristics: Stylar shelf of the upper molars reduced and stylar cusps reduced.
Genetic characteristics: 2n=14. In Caluromys the nucleolar organizing region (NOR) is located on a single pair of autosomes.
Parasites: Thatcher (2006) documents the Protozoa Trypanosoma for Brazil.
Paraguayan Species:
Caluromys lanatus - Brown-eared Woolly Opossum HBook 16
Subfamily Didelphinae Gray, 1821
Tribe Didelphini Gray, 1821
General characteristics: Five large opossums in four genera with total length >500mm. Pouch moderately to well-developed.
Didelphis Linnaeus, 1758: True Opossums
Five species, two of which occur in Paraguay. Synonyms adapted from Gardner (2007).
Synonyms:
Didelphis Linnaeus 1758:54. Type species Didelphis marsupialis Linnaeus (1758).
Didelphys Schreber 1777:532. Unjustified emendation.
Opossum Schmid 1818:115. In part. Didelphis marsupialis Linnaeus (1758).
Sarigua Muirhead 1819:429. In part.
Didelphus I. Geoffroy St.Hilaire 1831:139. Incorrect spelling.
Thylacotherium Lund 1839:233. Type species Thylacotherium ferox Lund (1839). Preoccupied by Thylacotherium Valenciennes 1838.
Micoureus Lesson 1842:186. In part.
Didelphus Lapham 1853:337. Incorrect spelling.
Gamba Liais 1872:329. In part.
Gambatherium Liais 1872:331. Replacement name for Thylacotherium Lund (1839).
Dasyurotherium Liais 1872:331. Replacement name for Thylacotherium Lund (1839).
Dimerodon Ameghino 1889:277. Type species Dimerodon mutilatus Ameghino (1889) by monotypy.
Leucodidelphis Ihering 1914:347. Type species Didelphis paraguayensis Oken (1816). Name not available. Proposed as subgenus of Didelphis.
Leucodidelphys Krumbiegel 1941:34. Unjustified emendation.
Leucodelphis Cabrera 1958:41. Incorrect spelling.
General characteristics: Two species of "typical" opossum with pointed snout, large, naked, rounded ears and long, prehensile tail furred at base and white towards the tip. This genus includes the largest of New World marsupials. Females have a well-developed pouch with 11 to 13 teats. Pelage is soft and dense with long, coarse guard hairs. Semi-arboreal in behaviour. Fossils from the middle Pleistocene of South America have been referred to this genus
Cranial characteristics: Well-developed sagittal crest in adults. Narrow braincase with interorbital width greater than the postorbital constriction.
Dental characteristics: Dental wear can be used for aging in this genus, see species accounts and (Fig 2).
Genetic characteristics: 2n=14. The nucleolar organizing region (NOR) is located on up to three pairs of autosomes.
Parasites: Linardi (2006) gives the following ectoparasites recorded in this genus from Brazil: Siphanaptera: Craneopsylla (Stephanocircidae); Ctenocephalides (Pulicidae); Polygenis and Rhopalopsyllus (Rhopalopsyllidae); Adoratopsylla (Ctenophthalmidae). Acari: Metastigmata Ixodes, Ornithodoros and Amblyomma (Ixodidae). Acari: Mesostigmata Ornithonyssus and Bdellonyssus (Macronyssidae); Androlaelaps, Laelaps and Gigantolaelaps (Laelapidae). Acari: Prostigmata Archemyobia (Myobiidae). Acari: Astigmata Didelphilichus (Atopomelidae). Thatcher (2006) documents the following endoparasites for Brazil: Protozoa Trypanosoma and Babesia. Trematoda Schistosoma (Schistosomatidae), Aphimerus (Opisthorchiidae), Rhopalias (Rhopaliasidae) and Paragonimus (Paragonimidae). Cestoda Mathevotaenia bivittata (Anoplocephalidae). Nematoda Dracunculus (Dracunculidae); Gongelonemoides (Gongylonematidae); Dipetalonema (Oncocercidae); Thominx (Trichuridae); Travassostrongylus and Viannaia (Trichostrongylidae). Acanthocephala Oligacanthorhynchus (Oligacanthorhynchidae).
Evolution: Formerly considered to be the most primitive of Didelphid lineages, Patton & Costa (2003) demonstrated that the genus is in fact young and speciated relatively recently.
Paraguayan Species:
Didelphis albiventris - White-eared Opossum
Didelphis aurita - Southern Black-eared Opossum
Chironectes Illiger, 1811: Water Opossum
This is a monotypic genus. Synonyms adapted from Gardner (2007).
Synonyms:
Latra Zimmermann 1780:317. In part. Incorrect spelling of Lutra. Not Lutra Linnaeus 1758.
Mustela Kerr 1792:172. In part. Not Mustela Linnaeus 1758.
Lutra Link 1795:172. Not Lutra Linnaeus 1758.
Didelphis G.Cuvier 1798:125. In part. Not Didelphis Linnaeus 1758.
Lutra G.Shaw 1800:447. Not Lutra Linnaeus 1758.
Mustela Turton 1800:58. In part. Not Mustela Linnaeus 1758.
Didelphis Daudin in Lacépède 1802:152. Not Didelphis Linnaeus 1758.
Chironectes Illiger 1811:76. Type species Lutra minima Zimmermann 1780 by monotypy.
Memina G.Fischer 1813:15. Nomen nudum.
Memina G.Fischer 1814:11. Type species Lutra memina Boddaert 1784 by monotypy.
Sarigua Muirhead 1819:429. In part.
Cheironectes Gray 1821:308. Incorrect spelling.
Cheronectis Fleming 1822:212. Incorrect spelling.
Gamba Liais 1872:329. Type species Gamba palmata Liais 1872 by designation.
Chironeytes Goeldi & Hagman 1904:100. Incorrect spelling.
General characteristics: Chironectes is the only truly aquatic opossum. The tail is longer than the head and body, naked and rounded in cross-section. It is not prehensile. The ears are fairly large, rounded and naked. The snout is long with stiff facial bristles. Further adaptations for an aquatic existence include the dense, short, waterproof pelage and the hindfeet webbed to the tips of the toes. The forefeet are unwebbed with long, dextrous fingers, clubbed at the end. A well-developed, backwards-opening pouch is present in both sexes and can be sealed whilst the animal is submerged. There are 4 or 5 mammae. Fossils from the Miocene of Argentina have been referred to this genus.
Cranial characteristics: Recalls that of Philander. Long rostrum, rounded braincase and broadly flared zygomatic arches. Palate with a single pair of maxillopalatine fenestrae opposite the molars. Small palatine fenestrae are absent.
Genetic characteristics: 2n=14. The nucleolar organizing region (NOR) is located on a single pair of autosomes.
Parasites: Linardi (2006) gives the following ectoparasites recorded in this genus from Brazil: Siphanaptera: Adoratopsylla (Ctenophthalmidae). Thatcher (2006) documents the following endoparasites for Brazil: Trematoda Rhopalias (Rhopaliasidae).
Paraguayan Species:
Chironectes minimus - Water Opossum
Lutreolina O. Thomas, 1910: Thick-tailed Opossum
This is a monotypic genus. Synonyms adapted from Gardner (2007).
Synonyms:
Didelphis Desmarest 1804:19. In part. Not Didelphis Linnaeus (1758).
Didelphys Olfers 1818:204. In part. Incorrect spelling.
Sarigua Muirhead 1819:429. In part.
Peramys Lesson 1842:261. In part.
Micoureus P.Gervais 1855:287. In part. Not Micoureus Lesson (1842).
Philander Gerard 1862:139. In part. Not Philander Brisson (1762).
Metachirus Hensel 1872:121. In part. Proposed as a subgenus of Didelphis. Not Metachirus Burmeister (1854).
Lutreolina O.Thomas 1910:247. Type species Didelphis crassicaudata Desmarest (1804) by monotypy.
General characteristics: Lutreolina bears a superficial resemblance to a small otter or weasel, with a thick, non-prehensile tail furred to approximately half its length. Pelage is uniformly-coloured, thick and dense but not waterproof. Living or freshly-killed specimens fluoresce red-orange under UV light. The snout is short and the ears small and rounded. The legs are short and strong. This species possesses only a slight pouch. Hallux is not fully opposable. Fossils from the Pliocene of Argentina have been referred to this genus
Cranial characteristics: Shows several adaptations related to a more carnivorous diet. Distinctive amongst Didelphids on account of the shortened rostrum, narrow braincase and zygomatic arches. The nasals taper evenly. Braincase narrower and longer than similar-sized species with widened temporal fossa and broader, more perpendicular coronoid processes. (Vieira & Astúa de Morães 2003)
Genetic characteristics: 2n=14. The nucleolar organizing region (NOR) is located on two pairs of autosomes.
Parasites: Linardi (2006) gives the following ectoparasites recorded in this genus from Brazil: Siphanaptera: Craneopsylla (Stephanocircidae); Polygenis (Rhopalopsyllidae); Ctenocephalides (Pulicidae); Adoratopsylla (Ctenophthalmidae). Acari: Metastigmata Ixodes (Ixodidae).
Paraguayan Species:
Lutreolina crassicaudata - Thick-tailed Opossum
Designed by Paul Smith 2006. This website is copyrighted by law.
Material contained herewith may not be used without the prior written permission of FAUNA Paraguay.
Photographs on this web-site and are used with permission.
Material contained herewith may not be used without the prior written permission of FAUNA Paraguay.
Photographs on this web-site and are used with permission.
Cáceres N 2006 - O Papel de Marsupiais na Dispersão de Sementes p255-270 in Cáceres NC, Monteiro-Filho ELA Os Marsupiais do Brasil:Biologia, Ecologia e Evolução - Editora UFMS, Campo Grande.
Contreras J & Silvera 1995 - Incorporación del pequeño marsupial Monodelphis scalops a la mastofauna del Paraguay - Notulas Faunisticas 70
Costa LP 2006 - Relações Filogenéticas Intergenéricas de Marsupiais Didelfídeos p343-361 in Cáceres NC, Monteiro-Filho ELA Os Marsupiais do Brasil:Biologia, Ecologia e Evolução - Editora UFMS, Campo Grande.
Costa LP, Leite YLR, Patton JL 2003 - Phylogeography and Systematic Notes on Two Species of Gracile Mouse Opossums, Genus Gracilinanus (Marsupialia: Didelphidae) from Brazil - Proceedings of the Biological Society of Washington 116: p275-292.
de la Sancha N, Solari S, Owen RD 2007 - First Records of Monodelphis kunsi Pine from Paraguay, with an Evaluation of its Distribution - Mastozoologia Neotropical 14: p241-247.
Diaz MM & Barquez RM 2002 - Los Mamíferos de Jujuy, Argentina - LOLA
Emmons LH & Feer F 1999 - Mamíferos de los Bosque Húmedos de América Tropical - FAN Bolivia
Gardner AL 2007 - Mammals of South America Volume 1: Marsupials, Xenarthrans, Shrews and Bats - University of Chicago Press.
Goin FJ 2003 - Early Marsupial Radiations in South America p30-42 in Jones M, Dickman C, Archer M Predators with Pouches: The Biology of Carnivorous Marsupials - CSIRO, Collingwood.
Hershkovitz P 1992 - The South American Gracile Mouse Opossums Genus Gracilinanus Gardner & Creighton 1989 (Marmosidae: Marsupialia): A Taxonomic Review with Notes on General Morphology and Relationships - Fieldiana Zoology Series 1441.
Hershkovitz P 1997 - Composition of the Family Didelphidae Gray 1821 (Didelphoidea: Marsupialia) with a Review of the Morphology and Behaviour of the Included Four-eyed Pouched Opossums of the Genus Philander Tiedemann 1808 - Fieldiana 86.
Hokoç JN, Aráujo Lima SM, Moraes AMM, Ahnelt P 2006 - A Visão em Marsupiais: Características e Evolucão p69-81 in Cáceres NC, Monteiro-Filho ELA Os Marsupiais do Brasil:Biologia, Ecologia e Evolução - Editora UFMS, Campo Grande.
Jansa SA, Forsman JF, Voss RS 2006 - Different Patterns of Selection on the Nuclear Genes IRBP and DMP-1 Affect the Efficiency but not the Outcome of Phylogeny Estimation for Didelphid Marsupials - Molecular Phylogenetics and Evolution 38: p363-380.
Jansa SA, Voss RS 2000 - Phylogenetic Studies on Didelphid Marsupials I: Introduction and Preliminary Results from Nuclear IRBP Gene Sequences - Journal of Mammalian Evolution 7: p43-77.
Jones M, Dickman C, Archer M eds. 2003 - Predators With Pouches: The Biology of Carnivorous Marsupials - CSIRO, Collingwood.
José H 2006 - Glândulas Paracloacais de Marsupiais p81-88 in Cáceres NC, Monteiro-Filho ELA Os Marsupiais do Brasil:Biologia, Ecologia e Evolução - Editora UFMS, Campo Grande.
Limardi PM 2006 - Os Ectoparasitos de Marsupiais Brasileiros p27-52 in Cáceres NC, Monteiro-Filho ELA Os Marsupiais do Brasil:Biologia, Ecologia e Evolução - Editora UFMS, Campo Grande.
Massoia E, Forasiepi A & Teta P 2000 - Los Marsupiales de la Argentina - LOLA.
Monteiro-Filho ELA, Cáceres NC 2006 - Biologia Reprodutiva de Fêmeas de Marsupiais Didelfídeos p99-110 in Cáceres NC, Monteiro-Filho ELA Os Marsupiais do Brasil:Biologia, Ecologia e Evolução - Editora UFMS, Campo Grande.
Nogueira JC 2006 - Morfologia do Sistema Genital Masculino de Marsupiais Brasileiros p111-130 in Cáceres NC, Monteiro-Filho ELA Os Marsupiais do Brasil:Biologia, Ecologia e Evolução - Editora UFMS, Campo Grande.
Nogueira JC, Castro ACS 2003 - Male Genital System of South American Didelphids p183-204 in Jones M, Dickman C, Archer M eds. 2003 - Predators With Pouches: The Biology of Carnivorous Marsupials - CSIRO, Collingwood.
O´Brien SJ, Menninger JC, Nash WC 2006 - Atlas of Mammalian Chromosomes - Wiley, New Jersey.
Patton JL, Costa LP 2003 - Molecular Phylogeography and Species Limits in Rainforest Didelphid Marsupials of South America p63-81 in Jones M, Dickman C, Archer M eds. 2003 - Predators With Pouches: The Biology of Carnivorous Marsupials - CSIRO, Collingwood.
Quadros J 2006 - A Identificacão de Marsupiais da Floresta Atlântica Basada na Análise Microscópica de Pêlos-guarda p29-36 in Cáceres NC, Monteiro-Filho ELA Os Marsupiais do Brasil:Biologia, Ecologia e Evolução - Editora UFMS, Campo Grande.
Redford K & Eisenberg JF 1992 - Mammals of the Neotropics Vol 2: The Southern Cone Chile, Argentina, Uruguay and Paraguay - University of Chicago Press.
Reig OA, Kirsch JAW, & Marshall LG 1985 - New Conclusions on the Relationships of the Opossum-like Marsupials with an Annotated Classification of the Didelphimorphia - Ameghiniana 21: p335-343.
Solari S 2003 - Diversity and Distribution of Thylamys (Didelphidae) in South America, with Emphasis on Species from the Western Side of the Andes p82-101 in Jones M, Dickman C, Archer M eds. 2003 - Predators With Pouches: The Biology of Carnivorous Marsupials - CSIRO, Collingwood.
Thatcher VE 2006 - Os Endoparasitos dos Marsupiais Brasileiros p53-68 in Cáceres NC, Monteiro-Filho ELA Os Marsupiais do Brasil:Biologia, Ecologia e Evolução - Editora UFMS, Campo Grande.
Tyndale-Briscoe H 2005 - Life of Marsupials - CSIRO, Collingwood.
Tyndale-Biscoe CH, MacKenzie RB 1976 - Reproduction in Didelphis marsupialis and D. albiventris in Colombia - Journal of Mammalogy 57: p249265.
Van Nievelt AFH, Smith KK 2006 - Tooth Eruption in Monodelphis domestica and its Significance for Phylogeny and Natural History - Journal of Mammalogy 86; p333-341.
Vieira EM, Astúa de Morães DA 2003 - Carnivory and Insectivory in Neotropical Marsupials p271-284 in Jones M, Dickman C, Archer M eds. 2003 - Predators With Pouches: The Biology of Carnivorous Marsupials - CSIRO, Collingwood.
Contreras J & Silvera 1995 - Incorporación del pequeño marsupial Monodelphis scalops a la mastofauna del Paraguay - Notulas Faunisticas 70
Costa LP 2006 - Relações Filogenéticas Intergenéricas de Marsupiais Didelfídeos p343-361 in Cáceres NC, Monteiro-Filho ELA Os Marsupiais do Brasil:Biologia, Ecologia e Evolução - Editora UFMS, Campo Grande.
Costa LP, Leite YLR, Patton JL 2003 - Phylogeography and Systematic Notes on Two Species of Gracile Mouse Opossums, Genus Gracilinanus (Marsupialia: Didelphidae) from Brazil - Proceedings of the Biological Society of Washington 116: p275-292.
de la Sancha N, Solari S, Owen RD 2007 - First Records of Monodelphis kunsi Pine from Paraguay, with an Evaluation of its Distribution - Mastozoologia Neotropical 14: p241-247.
Diaz MM & Barquez RM 2002 - Los Mamíferos de Jujuy, Argentina - LOLA
Emmons LH & Feer F 1999 - Mamíferos de los Bosque Húmedos de América Tropical - FAN Bolivia
Gardner AL 2007 - Mammals of South America Volume 1: Marsupials, Xenarthrans, Shrews and Bats - University of Chicago Press.
Goin FJ 2003 - Early Marsupial Radiations in South America p30-42 in Jones M, Dickman C, Archer M Predators with Pouches: The Biology of Carnivorous Marsupials - CSIRO, Collingwood.
Hershkovitz P 1992 - The South American Gracile Mouse Opossums Genus Gracilinanus Gardner & Creighton 1989 (Marmosidae: Marsupialia): A Taxonomic Review with Notes on General Morphology and Relationships - Fieldiana Zoology Series 1441.
Hershkovitz P 1997 - Composition of the Family Didelphidae Gray 1821 (Didelphoidea: Marsupialia) with a Review of the Morphology and Behaviour of the Included Four-eyed Pouched Opossums of the Genus Philander Tiedemann 1808 - Fieldiana 86.
Hokoç JN, Aráujo Lima SM, Moraes AMM, Ahnelt P 2006 - A Visão em Marsupiais: Características e Evolucão p69-81 in Cáceres NC, Monteiro-Filho ELA Os Marsupiais do Brasil:Biologia, Ecologia e Evolução - Editora UFMS, Campo Grande.
Jansa SA, Forsman JF, Voss RS 2006 - Different Patterns of Selection on the Nuclear Genes IRBP and DMP-1 Affect the Efficiency but not the Outcome of Phylogeny Estimation for Didelphid Marsupials - Molecular Phylogenetics and Evolution 38: p363-380.
Jansa SA, Voss RS 2000 - Phylogenetic Studies on Didelphid Marsupials I: Introduction and Preliminary Results from Nuclear IRBP Gene Sequences - Journal of Mammalian Evolution 7: p43-77.
Jones M, Dickman C, Archer M eds. 2003 - Predators With Pouches: The Biology of Carnivorous Marsupials - CSIRO, Collingwood.
José H 2006 - Glândulas Paracloacais de Marsupiais p81-88 in Cáceres NC, Monteiro-Filho ELA Os Marsupiais do Brasil:Biologia, Ecologia e Evolução - Editora UFMS, Campo Grande.
Limardi PM 2006 - Os Ectoparasitos de Marsupiais Brasileiros p27-52 in Cáceres NC, Monteiro-Filho ELA Os Marsupiais do Brasil:Biologia, Ecologia e Evolução - Editora UFMS, Campo Grande.
Massoia E, Forasiepi A & Teta P 2000 - Los Marsupiales de la Argentina - LOLA.
Monteiro-Filho ELA, Cáceres NC 2006 - Biologia Reprodutiva de Fêmeas de Marsupiais Didelfídeos p99-110 in Cáceres NC, Monteiro-Filho ELA Os Marsupiais do Brasil:Biologia, Ecologia e Evolução - Editora UFMS, Campo Grande.
Nogueira JC 2006 - Morfologia do Sistema Genital Masculino de Marsupiais Brasileiros p111-130 in Cáceres NC, Monteiro-Filho ELA Os Marsupiais do Brasil:Biologia, Ecologia e Evolução - Editora UFMS, Campo Grande.
Nogueira JC, Castro ACS 2003 - Male Genital System of South American Didelphids p183-204 in Jones M, Dickman C, Archer M eds. 2003 - Predators With Pouches: The Biology of Carnivorous Marsupials - CSIRO, Collingwood.
O´Brien SJ, Menninger JC, Nash WC 2006 - Atlas of Mammalian Chromosomes - Wiley, New Jersey.
Patton JL, Costa LP 2003 - Molecular Phylogeography and Species Limits in Rainforest Didelphid Marsupials of South America p63-81 in Jones M, Dickman C, Archer M eds. 2003 - Predators With Pouches: The Biology of Carnivorous Marsupials - CSIRO, Collingwood.
Quadros J 2006 - A Identificacão de Marsupiais da Floresta Atlântica Basada na Análise Microscópica de Pêlos-guarda p29-36 in Cáceres NC, Monteiro-Filho ELA Os Marsupiais do Brasil:Biologia, Ecologia e Evolução - Editora UFMS, Campo Grande.
Redford K & Eisenberg JF 1992 - Mammals of the Neotropics Vol 2: The Southern Cone Chile, Argentina, Uruguay and Paraguay - University of Chicago Press.
Reig OA, Kirsch JAW, & Marshall LG 1985 - New Conclusions on the Relationships of the Opossum-like Marsupials with an Annotated Classification of the Didelphimorphia - Ameghiniana 21: p335-343.
Solari S 2003 - Diversity and Distribution of Thylamys (Didelphidae) in South America, with Emphasis on Species from the Western Side of the Andes p82-101 in Jones M, Dickman C, Archer M eds. 2003 - Predators With Pouches: The Biology of Carnivorous Marsupials - CSIRO, Collingwood.
Thatcher VE 2006 - Os Endoparasitos dos Marsupiais Brasileiros p53-68 in Cáceres NC, Monteiro-Filho ELA Os Marsupiais do Brasil:Biologia, Ecologia e Evolução - Editora UFMS, Campo Grande.
Tyndale-Briscoe H 2005 - Life of Marsupials - CSIRO, Collingwood.
Tyndale-Biscoe CH, MacKenzie RB 1976 - Reproduction in Didelphis marsupialis and D. albiventris in Colombia - Journal of Mammalogy 57: p249265.
Van Nievelt AFH, Smith KK 2006 - Tooth Eruption in Monodelphis domestica and its Significance for Phylogeny and Natural History - Journal of Mammalogy 86; p333-341.
Vieira EM, Astúa de Morães DA 2003 - Carnivory and Insectivory in Neotropical Marsupials p271-284 in Jones M, Dickman C, Archer M eds. 2003 - Predators With Pouches: The Biology of Carnivorous Marsupials - CSIRO, Collingwood.
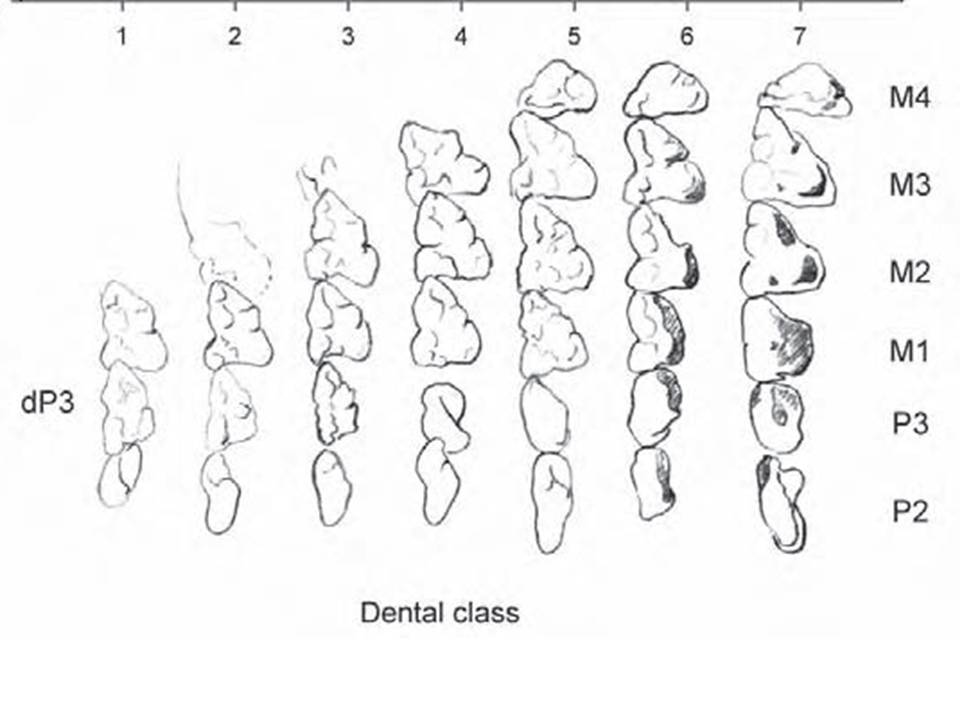
FIG 2: Tooth wear and age classes in Didelphis after Tyndale-Briscoe & McKenzie (1976).
Click to enlarge.
Click to enlarge.
FIG 3: Cranial bones of Didelphis sp. taken from Jolie M (1962) Chordate Morphology.
Click to enlarge.
Click to enlarge.