
Designed by Paul Smith 2006. This website is copyrighted by law.
Material contained herewith may not be used without the prior written permission of FAUNA Paraguay.
Photographs on this web-site were taken by Paul Smith, Hemme Batjes, Regis Nossent, Frank Fragano,
Alberto Esquivel, Arne Lesterhuis, José Luis Cartes, Rebecca Zarza and Hugo del Castillo and are used with their permission.
Tonatia bidens (Spix 1823)
TAXONOMY: Class Mammalia; Subclass Theria; Infraclass Metatheria; Order Chiroptera; Suborder Microchiroptera; Superfamily Noctilionoidea; Family Phyllostomidae, Subfamily Phyllostominae, Tribe Phyllostomini (López-Gonzalez 2005, Myers et al 2006, Hoffman et al 2008). There are two species in this genus, one of which occurs in Paraguay. Additional species assigned to this genus in the literature are now considered to be members of the genus Lophostoma (Lee et al 2002). The generic name Tonatia is of uncertain meaning. The species name bidens is Latin meaning double tooth referring to the two lower incisors, one of the distinguishing characteristics of this genus (Braun & Mares 1995).
The type specimen is an adult of unknown sex and without number in the Zoologisches Staats-Sammlung München, Germany with type locality "Fluvium St Francisci (=Rio São Francisco) Bahía, Brazil (Carter & Dolan 1978). The species is monotypic. References to this species in Central America and northern South America are referable to T.saurophila Koopman & Williams, 1951 (Williams et al 1995) a species initially described from fossil remains. Very little published data about the species today known as Tonatia bidens exists. Tonatia childreni was described based on a subadult specimen and was considered a synonym of this species by Williams et al (1995).
Synonyms adapted from Gardner (2007) and López-González (2005):
Vampyrus bidens Spix 1823:65. Type locality "Fluvium St Francisci (=Rio São Francisco) Bahía, Brazil.
[Tonatia] bidens Gray 1827:71. First use of current name combination.
Phyllostoma childreni Gray 1838:488 Type locality "South America".
Phyllostoma bidens Schinz 1844:236. Name combination.
Tylostoma bidens P.Gervais 1856:49 Name combination.
V[ampyrus]. (Tylostoma) bidens W.Peters 1856:304 Name combination.
Lophostoma bidens W.Peters 1865:509 Name combination.
Lophostoma bidens W.Peters 1865:509 Name combination.
Ph[yllostoma. (Tylostoma)]. Childreni W.Peters 1865:514 Name combination.
Tylostoma childreni Gray 1866:114 Name combination.
ENGLISH COMMON NAMES: Spix's Round-eared Bat (Gardner 2007), Greater Round-eared Bat (IUCN 2009).
SPANISH COMMON NAMES: Falso vampiro orejas redondas (Barquez et al1993), Murciélago de orejas redondas (Emmons 1999), Murciélago orejón grande (Ascorra et al 1991).
GUARANÍ COMMON NAMES: No known names.
DESCRIPTION: A small Phyllostomine with ears relatively short and broad when compared to Lophostoma (though large when compared to most other bats. Ears are round-tipped and do not extend beyond the nose when laid forwards, the inner margins of the proximal half also being bordered with lighter-coloured hairs. They are well-separated and not joined by a line of skin across the forehead. Tragus well-developed, antitragus clearly smaller than Lophostoma. Dorsal pelage varies from tawny-red (one specimen from Alto Paraguay TK 652259) through greyish to blackish-brown. Ventral pelage paler and greyer with buffy wash. Bases of hairs on the neck and behind the ears whitish, bases of other dorsal hairs dark brownish. Proximal half of the forearm, base of thumb and uppersides of feet furred. Nose leaf simple, broad at the base and narrowing suddenly from the middle to the somewhat blunt tip. Muzzle furred. Membranes dark brown and naked, save for a small strip of fur at the outer edge of the plagiopatagium. Upper lip smooth, lower lip with central wart surrounded by numerous small papillae. Tail small and protruding from the middle of the upper side of the uropatagium. Uropatagium broad and supported by small, weak calcars.
CRANIAL CHARACTERISTICS: Skull large and robust with broad, flat rostrum that lacks constriction in the orbital area. Low, broad braincase with upper edge evenly elevated from the front of the nasals and without a depression in the orbital region. Palate narrow. Sagittal crest low and slender. Lamboidal crest poorly developed. Zygomatic arches are slender, though appearing broad in lateral view, and have a medial constriction. Antero-medial foramen located between the upper incisors and incisive foramina. Secondary process on mastoid absent. Auditory bullae small, barely covering the middle of the cochlea. (Goodwin 1942, Williams et al 1995, Barquez et al 1999).
Species is sexually dimorphic in some measurements, as evidenced by the following from across the range (n=19-20 males, n=37-38 females) presented by Williams et al (1995): Greatest Length of Skull males 28.5mm (27.4-29.5mm), females 28.4mm (27.7-29.2mm); Condylobasal Length males 24.3mm (22.9-25.6mm), females 24mm (23.1-25.6mm); Zygomatic Width males 13.7mm (13.1-14.1mm), females 13.5mm (12.9-14.1mm); Interorbital Constriction males 5.9mm (5.6-6.1mm), females 5.9mm (5.5-6.2mm); Mastoid Width males 13.4mm (13-13.7mm), females 13.2mm (12.9-13.6mm); Braincase Width males 11.1mm (10.8-11.4mm), females 11.1mm (10.7-11.8mm); Width Across Canines males 6.1mm (5.5-6.3mm), females 6.1mm (5.8-6.4mm); Width Across Molars males 8.9mm (8.1-9.8mm), females 8.8mm (8.5-9.2mm).
Measurements of two male specimens from Paraguay from López-González (2005): Greatest Skull Length 26.6mm 27.4mm; Interorbital Constriction 5.8mm 5.7mm; Zygomatic Width 13.2mm 13.3mm; Mastoid Width 12.9mm 12.98mm; Width Across Molars 8.5mm 8.3mm; Width Across Canines 5.6mm 5.4mm.
Myers & Wetzel (1983) give the following measurements for a male and female respectively from Departamento Boquerón, Paraguay: Greatest Skull Length male 28.2mm; Condylobasal Length male 24.4mm; Zygomatic Width male 13.5mm; Mastoid Width male 13.2mm; Interorbital Constriction male 6mm; Length of Third Metacarpal male 48.3mm female 46.5mm; Width Across Upper Molars male 8.5mm; Width Across Upper Canines male 6.1mm female 5.6mm.
Barquez et al (1999) give the following measurements for individuals from Argentina (n=10 unless stated): Greatest Skull Length 27.9mm (+/- 0.46mm); Condylobasal Length 24.5mm (+/- 0.32mm); Zygomatic Width 13.6mm (+/- 0.22mm); Mastoid Width 13.3mm (+/- 0.13mm, n=7); Interorbital Constriction 7.2mm (+/- 0.4mm, n=5); Postorbital Constriction 5.7mm (+/- 0.12mm); Width Across Upper Molars 8.9mm (+/- 0.20mm); Width Across Upper Canines 5.9mm (+/- 0.28mm) Palatal Length 11.9mm (+/- 0.22mm); Length of Mandible 18mm (+/- 0.26mm, n=9); Width of Braincase 11.2mm (+/- 0.16mm).
Sanborn (1936) gives the following measurements for three specimens in the Chicago Field Museum: Greatest Length of Skull 27.8-28mm; Condylobasal Length 24-24.4mm; Length of Palate 12.6-13mm; Zygomatic Width 13.4-13.4mm; Interorbital Constriction 5.6-5.8mm; Mastoid Width 13-13.2mm;
Braincase Width 10.8-10.8mm; Width Across Canines 5.9-6mm; Width of Rostrum Across Premolars 5.5-5.6mm; Width Across Molars 7.9-8.4mm; Mandibular Length 17.2-17.6mm.
DENTAL CHARACTERISTICS: I2/2 C 1/1 P2/2 M3/3 = 32. Tooth rows converge slightly anteriorly. Inner incisors well-developed, in contact with each other, slightly procumbent and concave anteriorly, appearing slightly grooved. I2 is tiny and located within the cingulum of the canine. Lower incisors narrow, i1 weakly bilobed and higher than it is wide. Upper canines slightly curved. Lower canines either meet along posterior medial margin or are minutely separated. P1 is small and triangular in lateral view, with the anterior edge fitting completely in the canine cingulum. P2 is the highest in the toothrow. p1 and p3 are laterally compressed and unicuspidate. p2 reduced with tooth crown obscured by cingula of adjacent premolars. M1 and M2 are square with W-shape and well-developed cusps. M3 reduced with only two commissures, the parastyle being well-developed. Fourth commissure of m3 is reduced when compared to other molars. (Goodwin 1942, Williams et al 1995, Barquez et al 1999).
The following measurement from across the range (n=18 males, n=38 females) was presented by Williams et al (1995): Length of Upper Tooth Row males 9.9mm (9.5-10.2mm), females 9.8mm (9.4-10.2mm).
Measurements of two male specimens from Paraguay from López-González (2005): Length of Upper Tooth Row 9.5mm 9mm; Length of Lower Tooth Row 10.5mm 10.1mm.
Myers & Wetzel (1983) give the following measurements for a male and female respectively from Departamento Boquerón, Paraguay: Length of Upper Tooth Row male 10mm female 9.8mm.
Barquez et al (1999) give the following measurements for individuals from Argentina: Upper Tooth Row 10mm (+/- 0.31mm, n=10); Lower Tooth Row 11.3mm (+/- 0.21mm, n=6).
Sanborn (1936) gives the following measurements for three specimens in the Chicago Field Museum: Length of Upper Tooth Row 9.6-9.7mm; Length of Lower Tooth Row 10.8-10.9mm.
GENETIC CHARACTERISTICS 2n=16. FN=20. The X-chromosome is metacentric, the Y-chromosome acrocentric. (Baker & Hsu 1970, Baker 1973).
EXTERNAL MEASUREMENTS: A large bat but a small Phyllostomid. Species is sexually dimorphic in forearm measurements (n=19 males, n=38 females) from across the range presented by Williams et al (1995): FA: males 57.3mm (55.6-59.3mm), females 56.9mm (55.1-58.4mm).
Measurements of two male specimens from Paraguay from López-González (2005): TL 80mm 93mm; TA: 15mm 15mm; FT: 15mm 17mm; FA: 52mm 54mm; EA: 25mm 30; Length of Third Digit 45.4mm; WT: 26.9g 34g.
Myers & Wetzel (1983) give the following measurements for a male and female respectively from Departamento Boquerón, Paraguay: TL male 99mm; TA male 20mm; FT male 19mm; FA male 57.1mm female 54.9mm; EA male 27mm.
Barquez et al (1999) give the following measurements for individuals from Argentina (n=12 unless stated): TL 96.1mm (+/- 5.13mm); TA 16.5mm (+/- 2.36mm); FT 15.3mm (+/- 2.30mm); FA 57.6mm (+/- 1.16mm); EA 25.2mm (+/- 2.25mm); WT 32.7g (+/- 9.63mm, n=4).
Sanborn (1936) gives the following measurements for three specimens in the Chicago Field Museum: FA: 54.6-56.2mm; EA: 24.7-25.4mm; Length of Second Digit 34.4-36.6mm; Length of Third Metacarpal 45.6-48.5mm; Length of First Phalanx of Third Digit 18.7-19.6mm; Length of Second Phalanx of Third Digit 21.7-23.4mm; Length of Third Phalanx of Third Digit 16.5-18.5mm; Length of Fouth Metacarpal 46-49.9mm; Length of First Phalanx of Fourth Digit 14.5-16.1mm; Length of Second Phalanx of Fourth Digit 15.6-16.4mm; Length of Fifth Metacarpal 49-51.5mm; Length of First Phalanx of Fifth Digit 15.3-16.2mm; Length of Second Phalanx of Fifth Digit 13.7-15mm; Height of Nose Leaf 9.9-10.1mm; Width of Nose Leaf 6.4-6.5mm; Tibial Length 24.7-26.8mm; Calcar Length 17.9-19.1mm.
SIMILAR SPECIES: This is a small Phyllostominae (long ears, nose leaf well-developed with horseshoe shape enclosing the nostrils) with a short tail and round-tipped ears. Bats in the genus Lophostoma are extremely similar to Tonatia bidens and can be most easily separated on account of the naked or sparsely-furred muzzle and the fact that they roll their ears when handled. Both Tonatia and Lophostoma are unique amongst small Phyllostomids in having only one pair of lower incisors, the character being shared only by the much larger Chrotopterus auritus.
When compared to Lophostoma, Tonatia bidens has clearly separated ears, they are connected by a small band of skin in Lophostoma. Lophostoma brasiliense is much smaller with a forearm <45mm, whereas in this species the forearm is >50mm. Lophostoma silvicolum is of similar size to this species but has a variably conspicuous white throat patch that may be restricted to the chin or extend to the chest. Note also the unique shape of the tragus in L.silvicolum, being long, with three tooth-like projections near the base of the outer border, that of Tonatia bidens is shorter and with smooth, rounded edges (Genoways & Williams 1980). The antitragus is noticeably smaller in T.bidens than in L.silvicolum. Typically L.silvicolum has ear length >30mm whereas it is <30mm in this species.
Cranially the sagittal crest of L.silvicolum is more developed and the postorbital constriction of T.bidens is broader (>5mm in Tonatia, <5mm in L.silvicolum). Ratio of greatest skull length to postorbital constriction is <5.5mm in Tonatia, >5.9mm in L.silvicolum Note also the presence of antero-medial foramina behind the upper incisors in Tonatia, being absent in Lophostoma.
DISTRIBUTION: Locally distributed from northeast Brazil east of the Amazon south to Paraguay and northern Argentina. Literature references to the species presence in western and northern South America and Central America refer to T.saurophila.
In Brazil the species has been recorded in the following states: Bahía, Ceará, Espirito Santo, Minas Gerais, Mato Grosso do Sul, Mato Grosso, Pernambuco, Paran?, Rio de Janeiro, Santa Catarina and São Paulo (dos Reis et al 2007).
In Argentina it has been recorded in Provincias Jujuy and Misiones (Barquez et al 1993) though the original specimens were identified incorrectly as Lophostoma silvicolum (Barquez et al 1999). The species has not been recorded in Bolivia but possibly occurs there.
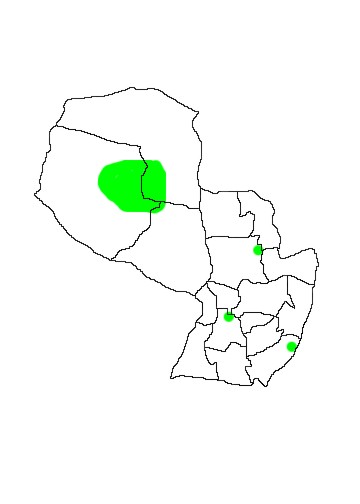
In Paraguay the species is known only from six diverse localities which suggest a more widespread distribution than currently known. Known localities are in Departamento Boquerón (km589 Ruta Trans-Chaco), Departamento Alto Paraguay (Estancia Tres Marías; 3 specimens TK 65259, 65260 and 65261), Departamento Concepción (Estancia San Luis), Departamento Itapúa (Arroyo San Rafael; UMMZ 125871), Departamento San Pedro (Yaguareté Forest; TK 56633) and Departamento Paraguarí (Sapucaí).
HABITAT: A low density habitat generalist able to utilise a range of both humid and dry habitats when sufficient food resources and roost sites are available.
Myers, White & Stallings (1983) captured specimens in Paraguay over an isolated pond in thorn (Chaco) scrub (Boquerón) and over a stream flowing through high tropical (Atlantic) forest (Itapúa). The species likely occurs widely through Paraguay in low densities in almost all natural habitats.
ALIMENTATION: Considered a foliage-gleaning insectivore by Willig (1985).
Foraging Behaviour and Diet Little known. Gardner (1977) lists fruit and insects, citing Ruschi (1953). Myers & Wetzel (1983) note that the stomach of a specimen in the High Chaco at Nueva Asunci?n, Departamento Boquerón contained large amounts of insect chitin and an unidentified whitish pulp. Barquez et al (1999) note that while observations of carnivorous feeding are rare, one specimen taken in Jujuy Argentina had a stomach filled with downy feathers, indicating recent consumption of a young bird.
REPRODUCTIVE BIOLOGY: Almost unknown. Virtually all published data refers to Tonatia saurophila.
Seasonality No data available for Paraguay.
Argentina A male with well-developed scrotal gonads was collected in Jujuy, Argentina in June (Barquez et al 1999).
Brazil Willig (1985) captured a pregnant female in the caatinga of northeast Brazil in September.
GENERAL BEHAVIOUR: No information.
Parasites Presley (2005) found 42 parasites on 3 specimens of this bat in Paraguay having a monoxenous spinturnicid (Periglischrus tonatii) and 70% of all Parichoronyssus crassipes detected in the country, despite the rarity of the host in the sample.
VOCALISATIONS: No information.
HUMAN IMPACT: None. This species is possibly naturally rare and rarely comes into contact with humans.
CONSERVATION STATUS: Globally considered to be Data Deficient by the IUCN, on account of ongoing taxonomic problems and an absence of recent data on range, status and ecological requirements. See http://www.iucnredlist.org/details/21983/0 for the latest assessment of the species. The species apparently occurs at naturally low population levels throughout its range. Following extensive sampling in Paraguay, Willig et al (2000) found this species to account for 0.19% of all bats caught in the Alto Chaco region (n=3989). The species might best be considered Data Deficient in Paraguay.
CITE AS: Smith P (2010) FAUNA Paraguay Online Handbook of Paraguayan Fauna Mammal Species Account 38 Tonatia bidens.
LAST UPDATED: 5 March 2010.
REFERENCES:
Ascorra CF, Wilson DE, Romo M 1991 - Lista Anotada de los Quirópteros del Parque Nacional Manu, Peru - Publicaciones del Museo de Historia Natural Universidad Nacional Mayor de San Marcos 42: p1-14.
Baker RJ 1973 - Comparative Cytogenetics of the New World Leaf-nosed Bats Phyllostomatidae - Periodic Biology 75: p37-45.
Baker RJ, Hsu TC 1970 - Further Studies on the Sex-chromosome Systems of the American Leaf-nosed Bats (Chiroptera: Phyllostomatidae) - Cytogenetics 9: p131-138.
Barquez RM, Giannini NP, Mares MA 1993 - Guide to the Bats of Argentina - Oklahoma Museum of Natural History.
Barquez RM, Mares MA, Braun JK 1999 - The Bats of Argentina - Museum of Texas Tech University Special Publications 42.
Braun JK, Mares MA 1995 - The Mammals of Argentina: An Etymology - Mastozoologia Neotropical 2: p173-206.
Carter DC, Dolan PG 1978 - Catalogue of Type Specimens of Neotropical Bats in Selected European Museums - Special Publications of the Museum of Texas Tech University 15.
Gardner AL 1977 - Feeding Habits p293-350 in Baker RJ, Jones JK, Carter DC Biology of Bats of the New World Family Phyllostomidae Part 2 - Texas Tech Museum Special Publications 13.
Gardner AL 2007 - Mammals of South America Vol 1: Marsupials, Xenarthrans, Bats and Shrews - University of Chicago Press, Chicago.
Gervais P 1856 - Deuxième Mémoire. Sur Quelques Points de l´Histoire Zoologique des Sarigues et Plus Particulièrement sur Leur Système Dentaire in Castelnau F de ed. Animaux Nouveau ou Rares Recueillis Pendant l´Expédition dans les Parties Centrales de l´Amérique du Sud, de Rio de Janeiro a Lima, et de Lima au Para; Executée par Ordre du Governement Français Pendant les Années 1843 a 1847, sous la Direction du Comte Francis de Castelnau. - P.Bertrand, Paris.
Goodwin GG 1942 - A Summary of Recognisable Species of Tonatia with Descriptions of Two New Species - Journal of Mammalogy 23: p204-209.
Gray JE 1827 - The Animal Kingdom Arranged in Conformity with its Organisation by the Baron Cuvier with Additional Descriptions of all the Species Hitherto Named and of Many Not Before Noticed by Edward Griffith....and Others - E.Griffith, London.
Gray JE 1838 - A Revision of the Genera of Bats (Vespertilionidae) and the Description of Some New Genera - Magazine Zoology and Botany 2: p483-505.
Gray JE 1838 - Revision of the Genera of Phyllostomidae or Leaf-nosed Bats - Proceedings of the Zoological Society of London 1849: p111-118.
Hoffman FG, Hoofer SR, Baker RJ 2008 - Molecular Dating of the Diversification of Phyllostominae Bats Based on Nuclear and Mitochondrial DNA Sequences - Molecular Phylogenetics and Evolution 49: p653-658.
Koopman KF, Williams EE 1951 - Fossil Chiroptera Collected by HE Anthony in Jamaica - American Museum Novitates 1519.
Lee TE, Hoofer SR, Van den Bussche RA 2002 - Molecular Phylogenetics and Taxonomic Revision of the Genus Tonatia (Chiroptera: Phyllostomidae) - Journal of Mammalogy 83: p49-57.
L?pez-Gonzalez C 2005 - Murciélagos del Paraguay - Biosfera Numero 9.
Miller GS 1907 - The Families and Genera of Bats - Bulletin USNM 57.
Myers P, Espinosa R, Parr CS, Jones T, Hammond GS, Dewey A 2006 - The Animal Diversity Web (online). Accessed December 2007.
Myers P, Wetzel RM 1983 - Systematics and Zoogeography of the Bats of the Chaco Boreal - Miscellaneous Publications of the Museum of Zoology, University of Michigan 165.
Myers P, White R, Stallings J 1983 - Additional Records of Bats From Paraguay - Journal of Mammalogy 64: p143-145.
Peters WCH 1856 - Über die Chiropterengattungen Mormops und Phyllostoma - Abhandl. K?nig Preuss. Akad. Wiss. Berlin 1857: p287-310.
Peters W 1865 - Über die zu den Vampyri Gehörigen Flederthiere und über die natürliche Stellung der Gattung - Antozous. Monatsber. K?nig. Preuss. Akad. Wiss. Berlin 1866: p503
Presley SJ 2005 - Ectoparasitic assemblages of Paraguayan bats: Ecological and Evolutionary Perspectives - Texas Tech University PhD Dissertation.
Reis dos NR, Peracchi AL, Pedro WA, Lima de IP 2007 - Morcegos do Brasil - Londrina, Brazil.
Ruschi A 1953 - Algumas observações sobre alimentação dos quiropteros, Phyllostomus hastatus hastatus (Pallas), Molossus rufus E.Geoffroy, Chrotopterus auritus australis (Thomas) e Noctilio leporinus leporinus (Linnaeus) - Boletim Museu Biologia Profesor Mello-Leitão, Sta Teresa 14: p1-5.
Sanborn CC 1936 - Records and Measurements of Neotropical Bats - Fieldiana Zoology 20: p93-106.
Schinz HR 1844 - Systematisches Verzeichniss alles bis jetzt Bekannten Säugethiere oder Synopsis Mammalium nach der Cuvier´schen System - Jent und Gassman, Solothurn.
Spix JB von 1823 - Simiarum et Vespertilionum Brasiliensum Species Novae: Ou Histoire Naturelle des Esepecies Nouvelles de Singes et Chauves-souris Observées et Recueilles Pendant le Voyage dans L´interieur de Bresil - Typis Francisi Serephici Hübschmanni, Monaco.
Williams SL, Willig MR, Reid FA 1995 - Review of the Tonatia bidens Complex (Mammalia: Chiroptera) with Descriptions of Two New Subspecies - Journal of Mammalogy 76: p612-626.
Willig MR 1985 - Reproductive Activity of Female Bats from Northeast Brazil - Bat Research News 26: p17-20.
Willig MA, Presley SJ, Owen RD, L?pez-Gonzalez C 2000 - Composition and Structure of Bat Assemblages in Paraguay: A Subtropical-Temperate Interface - Journal of Mammalogy 81: p386-401.
MAP 38:
Tonatia bidens